the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 15 Sep 2025
| 15 Sep 2025
Mapping benthic marine habitats featuring coralligenous bioconstructions: a new approach to support geobiological research
Giuseppe Maruca
Rocco Dominici
Gianpietro Imbrogno
Giovanni Vespasiano
Carmine Apollaro
Francesco Perri
Fabio Bruno
Antonio Lagudi
Umberto Severino
Valentina A. Bracchi
Daniela Basso
Emilio Cellini
Fabrizio Mauri
Antonietta Rosso
Rossana Sanfilippo
Adriano Guido
Seabed mapping represents a very useful tool for seascape characterization and benthic habitat study and requires advanced technologies for acquiring, processing, and interpreting remote data. Particularly, acoustic instruments, such as high-resolution swath bathymetry sounders (i.e., multibeam echosounder: MBES), allow us to recognize, identify, and map the extension of benthic habitats without applying invasive mechanical procedures. Bathymetry and backscatter (BS) data are crucial to perform modern habitat mapping. Although the acquisition and processing of bathymetric data follow standardized procedure (e.g., Hydrographic Organization guidelines), and recent studies have proposed recommendations for backscatter acquisition and processing, a broadly validated methodological approach, integrating geomorphometric analysis for benthic habitat mapping, is still lacking. In this work, a new approach for benthic habitat mapping, with a focus on coralligenous bioconstructions, was developed using the open-source software QGIS. This methodology, tested within the Isola Capo Rizzuto Marine Protected Area (Calabria, Italy), is designed to be freely reproducible by researchers working in the field of marine ecosystem monitoring and conservation. Through the proposed mapping procedure, it is possible to (i) identify benthic habitats in selected study areas by combining bathymetry and BS data with geomorphological indices performed in QGIS and (ii) quantitatively define the 2D and 3D distribution of coralligenous bioconstructions in terms of surface covered, thickness, and volume. Moreover, the statistical analysis of quantitative morphometric data allowed for comparison of geometric characteristics of different coralligenous morphotypes. The obtained results, combined with improvement of minimally invasive sampling and geobiological–geochemical characterization, can contribute to the development of protocols aimed at monitoring marine bioconstructed ecosystems, many of which are protected by national and international regulations due to their importance for Mediterranean biodiversity preservation, and plan actions for their protection and persistence.
- Article
(9454 KB) - Full-text XML
- BibTeX
- EndNote




Bioconstructions are geobiological bodies formed in situ by growth of skeletonized organisms and represent habitats that host a great variety of benthic species. They experience a wide array of dynamic phenomena, resulting from the balance between the action of habitat builders, dwelling organisms, and bio-eroders over decadal to millennial timescales. Along the Mediterranean continental shelf, the most conspicuous bioconstructed habitats are represented by coralligenous build-ups (Bracchi et al., 2015, 2017, 2022, 2025; Basso et al., 2022; Cipriani et al., 2023, 2024; Bazzicalupo et al., 2024, 2025; Sanfilippo et al., 2025), vermetid reefs (Picone and Chemello, 2023), sabellariid build-ups (Sanfilippo et al., 2019, 2022; Deias et al., 2023), and polychaete–bryozoan bioconstructions (Guido et al., 2013, 2016, 2017a, b, 2019a, b, 2022), whereas cold-water corals occur in deeper settings (Lo Iacono et al., 2018; Rueda et al., 2019; Foglini et al., 2019). Coralligenous is known as a biocenosis complex consisting of a hard biogenic substrate primarily generated by the superimposition of calcareous red algae able to form 3D structures, supporting a high biodiversity (e.g., Ballesteros, 2006; Bracchi et al., 2022; Rosso et al., 2023; Sciuto et al., 2023; Donato et al., 2024). Although recent studies have highlighted some terminological uncertainty in the definition of coralligenous habitat (e.g., Jardim et al., 2025, and references therein), within the geobiological literature the term coralligenous bioconstructions is widely and consistently adopted to indicate these biodiversity-rich, three-dimensional biogenic structures characterized by several layers of encrusting coralline algae (e.g., Ingrosso et al., 2018; Bracchi et al., 2017, 2022; Basso et al., 2022; Cipriani et al., 2023, 2024; Ferrigno et al., 2024).
Pérès and Picard (1964) and Pérès (1982) identified coralligenous as the ecological climax stage for the Mediterranean circalittoral zone, with some bioconstructions also occurring in dim-light very shallow settings (Ballesteros, 2006; Bracchi et al., 2016; Basso et al., 2022). Coralligenous produces various morphotypes on the seafloor and plays a key role in the formation and transformation of seascape over geological time (Bracchi et al., 2017; Marchese et al., 2020). Architecture and morphology are mainly influenced by biological carbonate production, which responds to different factors, like physiography, oceanography, terrigenous supply, and climate (Schlager, 1991, 1993; Betzler et al., 1977; Bracchi et al., 2017). Based upon the nature of the substrates, coralligenous morphotypes have been categorized in two main groups: (i) banks, flat frameworks mainly built on horizontal substrata and, and (ii) rims, structures on submarine vertical cliffs or close to the entrance of submarine caves (Pérès and Picard, 1964; Laborel, 1987; Ballesteros, 2006; Bracchi et al., 2017; Marchese et al., 2020; Gerovasileiou and Bianchi, 2021). Moreover, Bracchi et al. (2017) introduced a new classification for coralligenous morphotypes on subhorizontal substrate using a shape geometry descriptor in order to obtain a more objective description of these morphologies, classified into the following: (i) tabular banks, i.e., large tabular structures with a significant lateral continuity that completely cover the seafloor, forming an extensive habitat; (ii) discrete reliefs, i.e., smaller, distinct structures often arranged in clusters that do not fully cover the seafloor, leaving patches of sediment between them; and (iii) hybrid banks, a category grouping intermediate morphologies between tabular banks and discrete reliefs. These structures can coalesce into a larger formation, resembling tabular banks, while still maintaining individual characteristics. Hybrid banks often occur alongside other habitats, and their distribution is influenced by local sediment and hydrodynamic conditions (Bracchi et al., 2017).
Although coralligenous bioconstructions occur along almost the entire Mediterranean continental shelf, they have been mapped only in a few areas and their distribution is still underestimated (De Falco et al., 2010, 2022; Innangi et al., 2024). In addition, as known hot spot of biodiversity, along with its low accretion rate of 0.06–0.27 mm yr−1 and its sensitivity to natural and anthropogenic impacts (Di Geronimo et al., 2001; Bertolino et al., 2014; Basso et al., 2022; Cipriani et al., 2023, 2024), coralligenous is acknowledged as a priority habitat for protection under the EU Habitats Directive, is part of the Natura 2000 network (92/43/CE), and is subject to specific conservation plans within the framework of the Barcelona Convention (UNEP-MAP-RAC/SPA, 2008; UNEP-MAP-RAC/SPA, 2017). Moreover, together with other vulnerable settings (e.g., cold-water corals), coralligenous is monitored under the Marine Strategy Framework Directive (MSFD, EC, 2008; SNPA, 2024). As a result, nondestructive methods have been developed to assess the health status and ecological quality of this habitat (Bracchi et al., 2022). For all these reasons, seabed mapping can provide a very useful tool for seascape characterization and mapping of coralligenous and other vulnerable habitats (Chiocci et al., 2021). In particular, acoustic instruments, such as high-resolution swath bathymetry sounders, side scan sonars, and acoustic profiling instruments, enable the quick detection and identification of benthic habitats and thus the mapping of their extension without any direct contact that might represent a threat for these vulnerable ecosystems (Bracchi et al., 2017; Chiocci et al., 2021).
Several studies have demonstrated that such technologies, especially when combined with backscatter (BS) data and geometric descriptors, significantly enhance the study of seafloor properties and the discrimination of benthic habitats, such as coral reefs, improving the understanding of their spatial distribution and ecological significance (Fonseca and Mayer, 2007; Lecours et al., 2015, 2016; Brown et al., 2012; Lamarche and Lurton, 2018; Abdullah et al., 2024).
In this work, a semiautomated GIS-based approach for benthic habitat mapping was proposed and tested in shallow coastal waters, off Capo Bianco, within the Isola Capo Rizzuto Marine Protected Area (Crotone, southern Italy). The method combines high-resolution bathymetric and BS data obtained through multibeam echosounder (MBES) surveys and geomorphological and geomorphometric indices in order to develop innovative approaches for eco-geomorphological and geobiological characterization of the seafloor. The benthic habitat mapping proposed here has proven to be capable not only of identifying marine bioconstructions, but also of quantitatively defining their spatial and three-dimensional distribution in terms of area, volume, and height relative to the substrate from which they arise. For these reasons, the procedure represents a powerful tool for accurately delineating the extension of the bioconstructions and evaluating their evolution over time in response to natural and/or anthropogenic changes. Furthermore, the combination of this mapping approach with minimally invasive sampling systems and geobiological–geochemical characterization of marine bioconstructions may represent a potent tool for monitoring these delicate habitats.
High-resolution acoustic data of the study area offshore of Capo Bianco were collected during several MBES surveys (Fig. 1) performed between February and July 2024 as part of the project “Tech4You PP2.3.1: Development of tools and applications for integrated marine communities and substrates monitoring; Action 1: Development of hardware and software systems for three-dimensional detection, sampling and mapping of underwater environments”, an implementation of the previous bathymetric and backscatter data acquisition and elaboration of CRSM-ARPACAL.
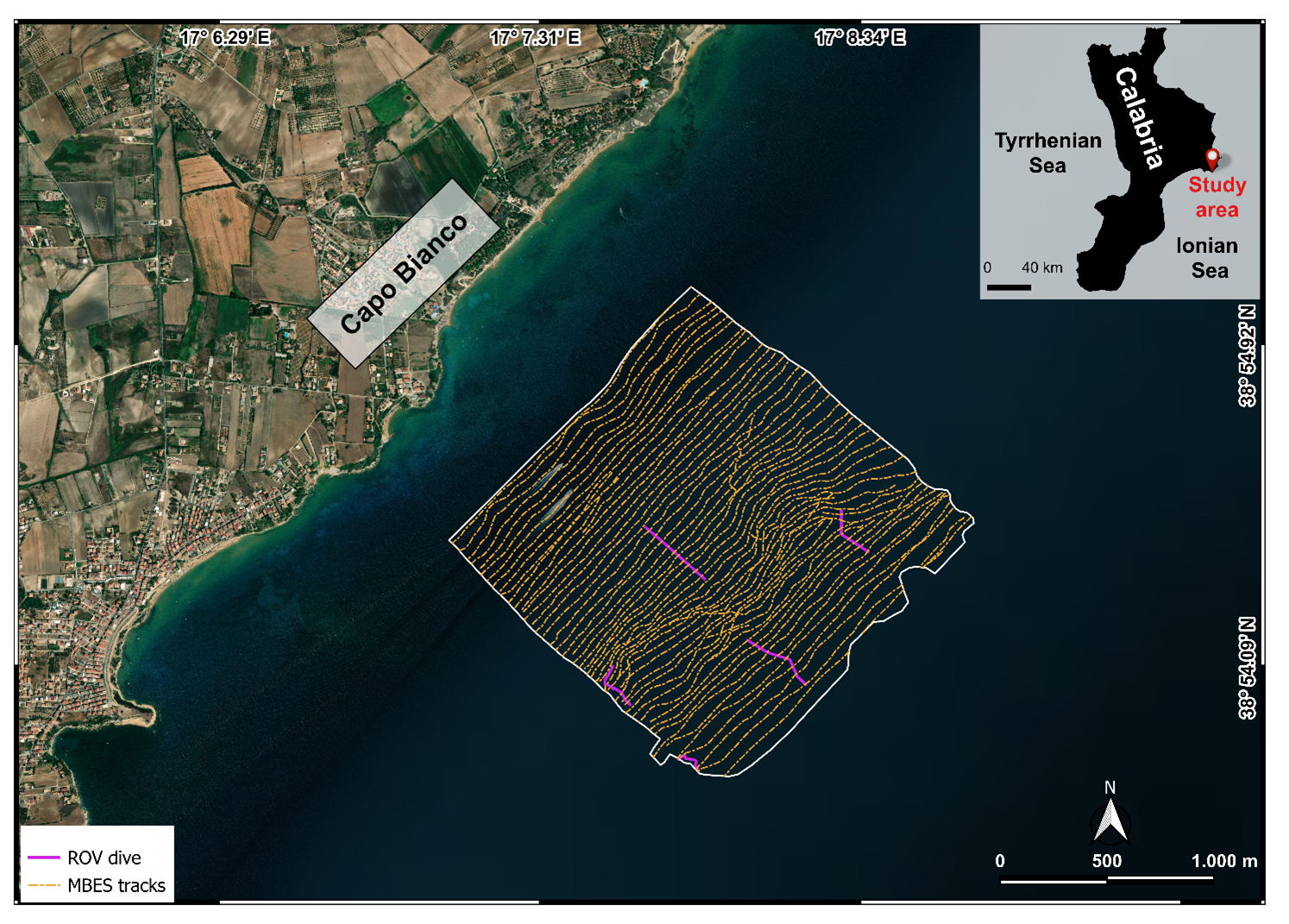
Figure 1Study area off Capo Bianco (Calabria, Italy) and location of the MBES tracks and ROV video transect (basemap from Esri World Imagery).
The approach proposed for benthic habitat mapping and defining of spatial and three-dimensional distribution of coralligenous bioconstruction is shown in Fig. 2. In particular, mapping operations were conducted using QGIS 3.34.9 “Prizren”. The most representative morphological indices, represented by slope and seafloor roughness, were extracted from the digital terrain model (DTM). Due to the large amount of data resulting from the need to obtain a high-resolution mapping of benthic habitats, backscatter and bathymetry values, together with geomorphological–geomorphometric indices, were imported and queried into PostgreSQL, an open-source and free relational database management system (RDBMS) capable of executing queries in Structured Query Language (SQL).
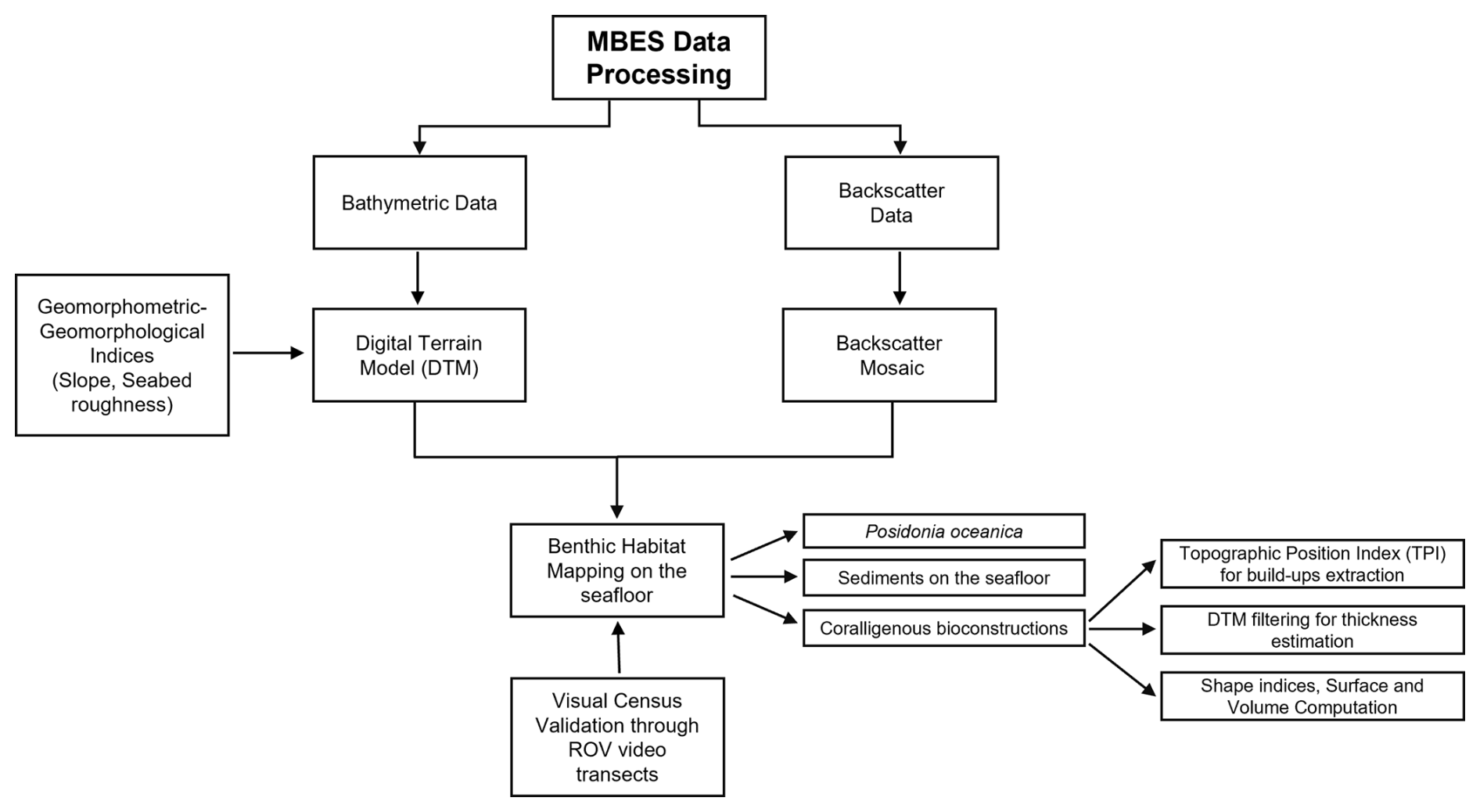
Figure 2Conceptual model of the workflow for the development of the proposed benthic habitat mapping approach.
Once the spatial extension and distribution of the benthic habitat had been defined by a combination of bathymetric, backscatter, slope, and seafloor roughness data, the extraction of coralligenous build-ups was performed using the topographic position index (TPI), according to Marchese et al. (2020). Moreover, area, shape index (SI), maximum diameter (Dmax) thickness, and volume were calculated for each extracted polygon. Finally, the benthic habitat mapping model was ground-truthed by visual analysis of remotely operated vehicle (ROV) video transects performed along specific paths within the study area. The underwater video surveys were obtained using a VideoRay Defender equipped with a functional prototype of the optical module dedicated to mapping, comprising a stereo camera, a high-resolution camera, and a lightning system (Severino et al., 2023). Both cameras have been meticulously calibrated to correct for optical distortions, ensuring accurate and reliable data acquisition. The selected cameras were the GoPro Hero 9 Black, serving as the high-resolution camera, and the Stereolabs ZED2i, serving as the stereo camera. The GoPro Hero 9 Black is a small-sized action camera with a 26.3 MP CMOS sensor capable of acquiring videos at a resolution of 5120×2880 at 30 fps, digital stabilization, and a horizontal field of view up to 128°. The ZED2i is a stereo camera with dual 4 MP sensors of 2 µm pixel size, a depth range between 0.3 to 20 m, a video resolution of 2208×1242 at 15 fps, and a horizontal field of view of 110°. The stereo camera communicates with the surface control unit by means of a single-board microcomputer, a NVIDIA Jetson Nano, which supports the CUDA architecture for parallel elaboration. The GoPro Hero 9 Black features Bluetooth low-energy (BLE) and Wi-Fi communication capabilities. The acquisition parameters for both cameras can be configured via the enclosure using a custom user interface accessible on the surface computer.
2.1 Bathymetric and backscatter data
MBES surveys have been carried out using a pole-mounted, Norbit iWBMS Long-Range Turnkey Multibeam Sonar System integrated with GNSS/INS (Applanix OceanMaster), operating with real-time kinematic (RTK) corrections, ensuring high positioning accuracy during the surveys. Data were collected in 59 tracks with a swath overlap of 20 %–40 % performed at an average speed of 2.315 m s−1. A total of three sound velocity profiles per day were collected before starting the acquisition using a sound velocity profiler: Valeport miniSVP. Considering the absence of freshwater inputs and the relative stability of the water column across the depth range, this was deemed sufficient to ensure reliable sound speed correction.
The MBES survey provided both bathymetry and BS data. The processing of MBES bathymetric data was performed using QPS Qimera and included corrections for tide, heading, heave, pitch, and roll. The correction of sound velocity was carried out using profiles obtained with the Valeport miniSVP. Subsequently, the soundings underwent manual cleaning to remove spikes. The bathymetric dataset was exported as a 32-bit raster file with a cell size of 0.05 m, obtained through interpolation of soundings using a CUBE surface model.
Backscatter data were processed using QPS Fledermaus, based on time series data and applying standard corrections for sonar configuration (e.g., source level, beam pattern, receiver gain) and environmental factors (e.g., absorption, slant range, footprint geometry). The processing was performed according to the general principles outlined in the Backscatter Working Group guidelines (Lurton et al., 2015), which provide detailed recommendations for the acquisition, correction, calibration, and processing of MBES backscatter data. The final output, exported as an 8-bit raster file with a 0.05 m cell size, was used to extract morphological and acoustic patterns of the seafloor.
2.2 Geomorphological–geomorphometric indices
Geomorphologic and geomorphometric indices were obtained using SAGA (System for Automated Geoscientific Analysis; Conrad et al., 2015) Next Gen Provider and GDAL plugins. In particular, the slope, expressed in degrees, was calculated using the dedicated function implemented in the GDAL plugin using a ratio of vertical units to horizontal of 1.0 and applying the Zevenbergen–Thorne formula instead of the Horn formula. Indeed, the Zevenbergen–Thorne method (Zevenbergen and Thorne, 1987), which considers a second-order finite difference, is more dedicated to geomorphological applications as it uses a particular weighting scheme that emphasizes changes in curvature and terrain shape. Seabed roughness was assessed using the terrain roughness index (TRI), which provides a quantitative measure of terrain heterogeneity (Riley et al., 1999). In particular, TRI values close to 0 indicate fairly regular and uniform surfaces, moderate TRI values correspond to more pronounced irregularities, and high TRI values identify rugged morphologies and/or complex structures on the seafloor. TRI was calculated using the SAGA module “Terrain Roughness Index” with the following settings: a circle as a search mode, a search radius of 0.5 m, a Gaussian weighting function, a value of 3.00 for the power, and a bandwidth of 75.00. The values of these parameters were selected though a trial-and-error method in order to best highlight the heterogeneity of the seabed.
2.3 Topographic position index
The topographic position index (TPI) was calculated at the finest possible scale (minimum radius: 1.00 m; maximum radius: 5.00 m) according to the DTM resolution and using a power of 3.00 and a bandwidth of 150.00. TPI is a morphometric parameter based on neighboring areas useful in DTM analysis (Wilson and Gallant, 2000). Specifically, positive TPI values indicate areas that are higher than the average of their surroundings, TPI values near zero correspond to flat areas or regions with a constant slope, and negative TPI values represent areas lower than their surroundings. In order to facilitate the extraction of coralligenous build-ups from surrounding seafloor and reduce the occurrence of artifacts, a TPI threshold of 0.2 was used and all the grid cells below this value were not considered to be coralligenous bioconstructions. The TPI scale (1.00–5.00 m) and value (0.2) were chosen through a trial-and-error approach in order to preserve the high resolution of the extraction, which is crucial for accurate volume computation.
2.4 DTM filtering
TPI parameters extracted the distribution of the coralligenous build-ups with high resolution in terms of perimeter boundaries. The thickness calculation for each coralligenous build-up was developed by the creation of a “reference surface” (without build-ups) using the SAGA DTM Filter (slope-based) tool implemented in QGIS 3.34.9. This tool uses concepts as described by Vosselman (2000) and can be used to filter a DTM, categorizing its cells into ground and non-ground (object) cells. A cell is considered ground if there is no other cell within the kernel radius where the height difference exceeds the allowed maximum terrain slope at the distance between the two cells. The thickness estimation of each coralligenous build-up was obtained by subtracting the average depth of each polygon extracted using TPI from the average depth value of the reference surface at that specific zone.
After estimating the height of each build-up relative to the seabed on which it developed, the shape index (SI – McGarigal and Marks, 1995) was calculated using the module “Polygon Shape Indices” in SAGA in order to describe a seafloor landscape characterized by distinct coralligenous morphotypes. Finally, the covered surface and volume of each polygon were calculated using a vector field operation implemented into QGIS.
The study area, located offshore of Capo Bianco (Isola Capo Rizzuto, Calabria, Italy), belongs to the Crotone Basin (CB) (Fig. 3), the widest Neogene basin of the Calabria region, partly exposed along the Ionian coast and partly documented offshore. It represents a segment of the Ionian forearc basin on the inner portion of the Calabrian accretionary wedge (Cosentino et al., 1989; Cavazza et al., 1997; Bonardi et al., 2001; Minelli and Faccenna, 2010). The basin infill is structured into several distinct tectono-stratigraphic sequences, which reflect an extensional to transtensional tectonic regime, occasionally interrupted by transpressional to compressional events (Malinverno and Ryan, 1986; Faccenna et al., 2001; Reitz and Seeber, 2012; Zecchin et al., 2012; Massari and Prosser, 2013; Milia and Torrente, 2014).
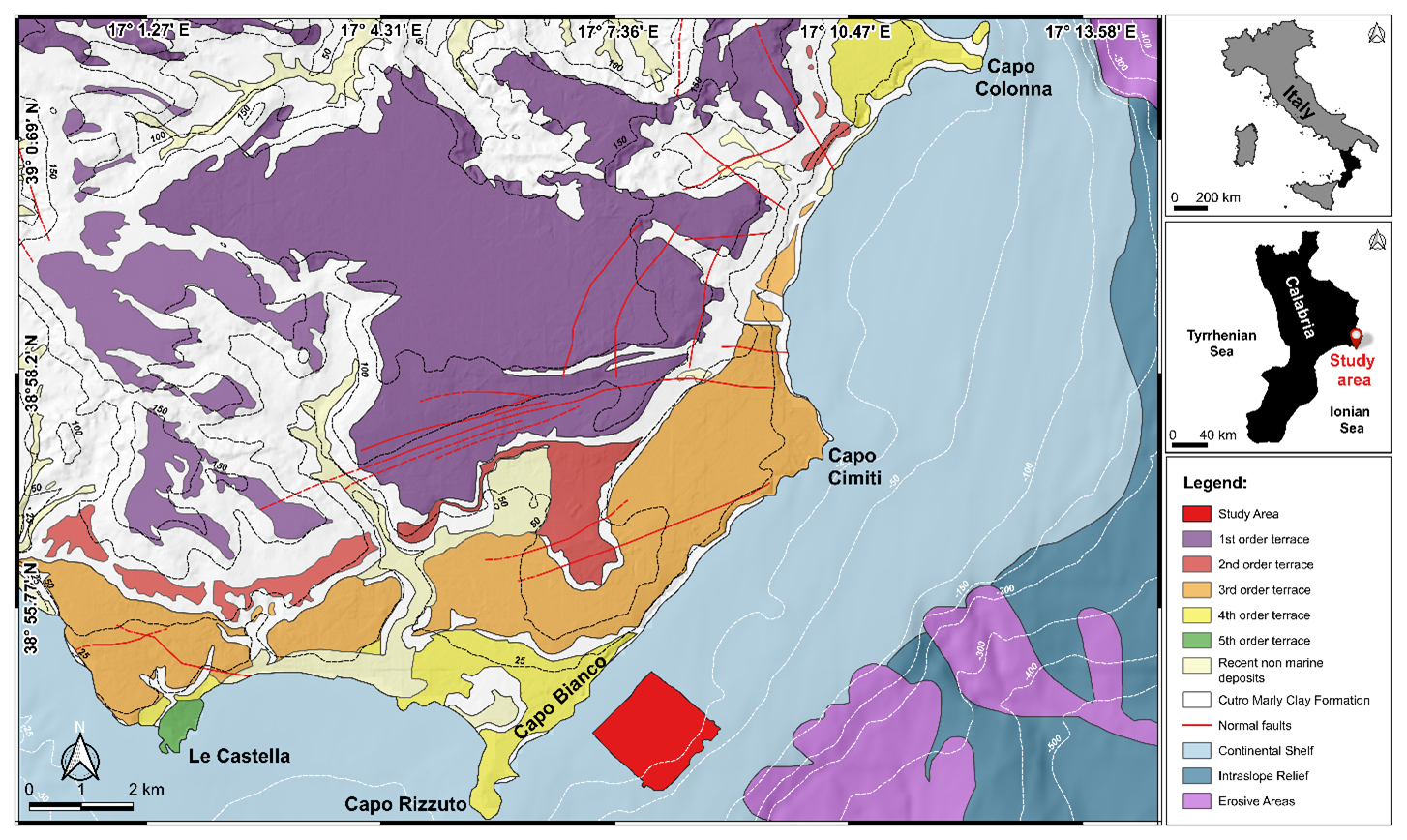
Figure 3Conflated geological map of the Crotone peninsula, with the indication of the five orders of terraces (modified from Bracchi et al., 2014), and physiographic domains identified offshore of the area in the framework of the MaGIC project (modified from Chiocci et al., 2021).
Since the mid-Pleistocene, a significant uplift (0.70–1.25 m kyr−1; Zecchin et al., 2004), combined with glacio-eustatic sea level fluctuations, has led to the formation in the Crotone Peninsula of five orders of marine terraces (Palmentola et al., 1990; Westaway, 1993; Westaway and Bridgland, 2007; Santoro et al., 2009; Faccenna et al., 2011; Bracchi et al., 2014; Santagati et al., 2024), which unconformably overlie the Piacenzian–Calabrian marly clays of the Cutro Formation (Zecchin et al., 2004).
The Cutro terrace (first-order terrace), ascribed to MIS 7 (Marine Isotope Stage 7) by Zecchin and Caffau (2011), is a mixed marine to continental terrace, consisting of the products of carbonate (algal build-ups and biocalcarenite passing into shoreface and foreshore deposits) to siliciclastic (shoreface, fluvial channel fill, lagoon–estuarine, and lacustrine deposits) sequences (Zecchin and Caffau, 2011).
The second order (MIS 5e), represented by the Campolongo–La Mazzotta terrace, is characterized by bioclastic and siliciclastic sandstones, with local bioclastic deposits and algal patch reefs (Mauz and Hassler, 2000; Zecchin and Caffau, 2011).
The Le Castella–Capo Cimiti terrace (third-order terrace), probably associated with MIS 5c (Mauz and Hassler, 2000), shows extensive algal reefs and shoreface deposits, with elevation variations due to normal fault displacement (Zecchin et al., 2004; Nalin et al., 2012).
The Capo Colonna marine terrace (fourth-order terrace), correlated with MIS 5.3 (Palmentola et al., 1990; Zecchin et al., 2004, 2009) or MIS 5.1 (ca. 80 ka; Gliozzi, 1987; Belluomini et al., 1988; Nalin et al., 2006; Nalin and Massari, 2009), consists of a planar surface with a sedimentary cover overlaid by a wedge of colluvium tapering (Bracchi et al., 2014).
The Le Castella marine terrace (fifth-order terrace) records an unconformity-bounded transgressive–regressive cycle (Nalin et al., 2007; Nalin and Massari, 2009; Zecchin et al., 2010; Bracchi et al., 2014, 2016), with two different facies for coralline algal build-ups and associated bioclastic deposits in the lower portion (Zecchin et al., 2004; Zecchin and Caffau, 2011). The age of these deposits remains debated, as they have been correlated with MIS 5.3 (Gliozzi, 1987), MIS 5.1 (Palmentola et al., 1990) and MIS 3 (Zecchin et al., 2004; Mauz and Hassler, 2000; Santagati et al., 2024).
The marine terraces exposed in the emerged portion near the study area demonstrated extensive carbonate production due to the development of algal bioconstruction throughout the Late Pleistocene. This production also appears to currently affect the seafloor. However, although the onshore portion of the CB has been well studied, its offshore extension is still less known (Pepe et al., 2010). Nevertheless, data from the MaGIC project related to Sheet 39 “Crotone” covered a vast area extending from the Neto Submarine Canyon to the Capo Rizzuto Swell. In this section, the continental shelf reaches up to 7 km wide, with the shelf break located at depths of 80–120 m. The slope encompasses the southern portion of the Neto Canyon headwall and the Esaro Canyon along with its tributaries. The average continental slope gradient is less than 5° and is characterized by an undulating morphology including the Luna and the Capo Rizzuto Swell. The southern section of the sheet covers the offshore extension of the Crotone forearc basin (Chiocci et al., 2021). This work aims to enhance the understanding of the Crotone Basin offshore features, with a focus on underwater bioconstructed habitats.
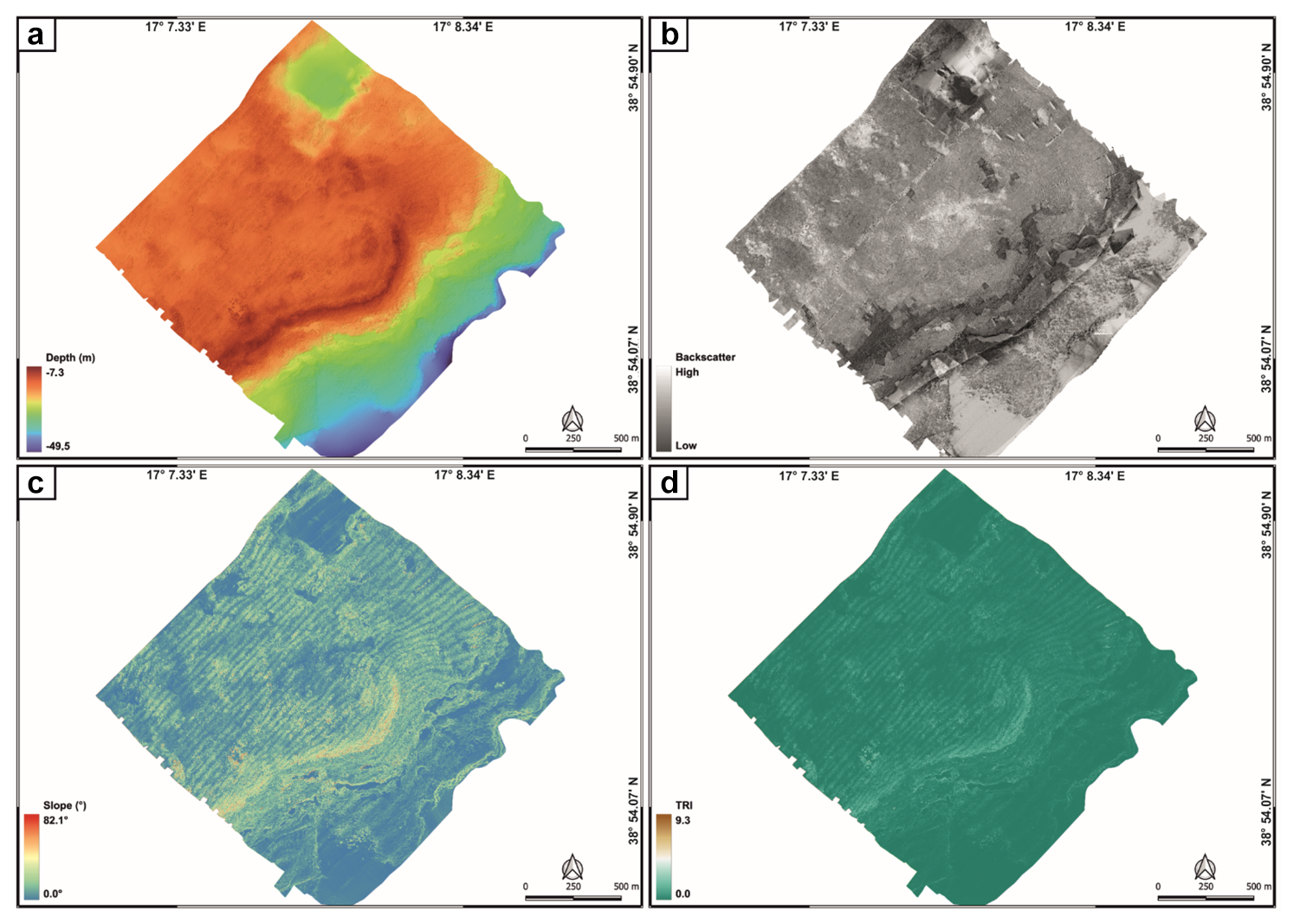
Figure 4Geomorphological characteristics of the study area expressed through processed bathymetric data (a), backscatter data (b), and geomorphometric indices, like slope (c) and terrain roughness index (d).
4.1 Morpho-acoustic characteristics of the seafloor
The comparison between bathymetric (Fig. 4a) and backscatter (Fig. 4b) data with those related to slope (Fig. 4c) and seafloor roughness (Fig. 4d) allowed for the definition of the morphological and morpho-acoustic characteristics of the study area off Capo Bianco (Calabria, Italy) and the identification of the benthic habitats. In particular, bathymetric data revealed a seafloor with depths ranging from −7.3 to −49.5 m (Fig. 4a). The transition towards the deeper areas is not gradual but shows an evident break in slope (starting from about −15 m depth), especially in the central zone of the study area. The shallower portion is characterized by widespread irregularities, while the deeper areas appear generally more regular, with less pronounced variations. Slope analysis (Fig. 4c) reveals maximum values (up to about 80°) along the break in slope, highlighting a steep and well-defined margin. The surrounding areas show lower slopes, with scattered peaks associated with seafloor irregularities. The terrain ruggedness index showed (i) a higher roughness along the break in slope (where the highest TRI values were recorded) and in its immediate vicinity and (ii) the presence of scattered roughness associated with irregularities on the seafloor (Fig. 4d).
Combining bathymetric and backscatter (Fig. 4b) data with slope and seafloor roughness values, different morpho-acoustic features were identified (Fig. 5).
-
Posidonia oceanica meadows, characterized by an intermittent speckled fabric of moderate backscatter. Posidonia covers seabed areas characterized by low slopes and slight roughness, spanning a depth range from about −6 to −25 m. In the depth range from −1 to −25 m, analysis of ROV video transects showed that the Posidonia meadow forms a mosaic with the coralligenous habitat.
-
Banks of coralligenous, characterized by a complex fabric of moderate to low backscatter. They covered areas characterized by moderate to high slopes and medium to high roughness, spanning a depth range from about −15 to −25 m,
-
Discrete coralligenous build-ups surrounded by medium to coarse sediment and maerl are characterized by a dotted pattern of moderate backscatter. They covered areas characterized by low slopes and medium roughness and occupy the area between the end of the banks and the final depth of the MBES survey, at approximately −40 m depth.
-
Fine to medium sediment, characterized by homogeneous patterns of medium to high backscatter. It covers scattered portions throughout the study area at various depths and is characterized by very low TRI values.

Figure 5Morpho-acoustic features identified by bathymetric and BS data, together with ROV video interpretation. The white scale bar is 20 m.
The combination of the various morpho-acoustic features enabled the identification of four main benthic habitats (Fig. 6): (i) Posidonia oceanica meadows, (ii) mosaic of coralligenous and Posidonia, (iii) coralligenous sensu stricto (i.e., bioconstructions that are not spatially intermixed with Posidonia oceanica), and (iv) fine to medium sediment.
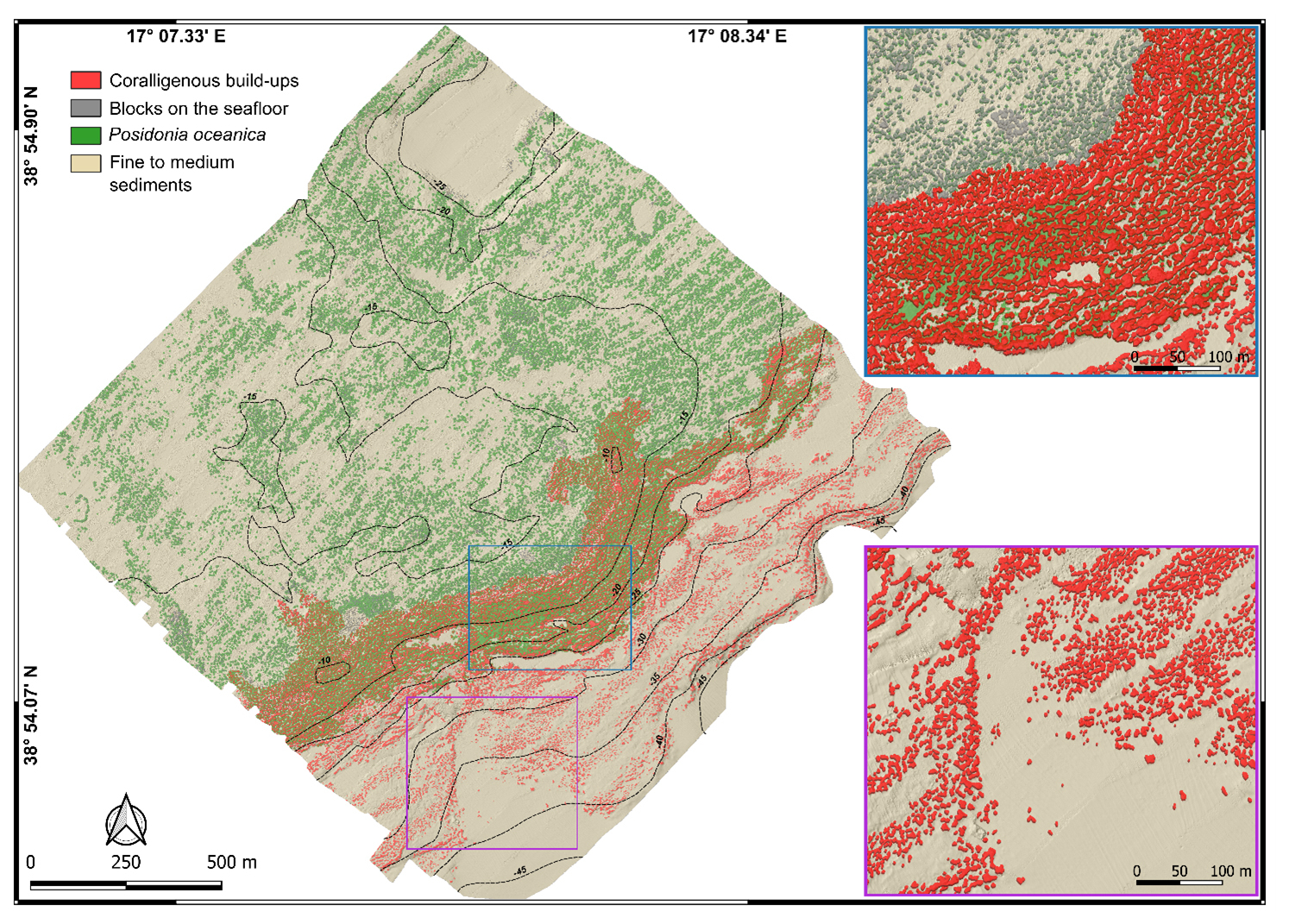
Figure 6Mapping model of the underwater benthic habitats in the study area off Capo Bianco (Calabria, Italy). Note, in the blue and purple boxes, two magnifications of representative areas of the model where coralligenous bioconstructions and rocky blocks on the seabed are depicted in 2.5D.
The Posidonia habitat, testified by its typical BS signal (intermittent speckled fabric of moderate backscatter), dominates in shallow areas (down to about −15 m depth), where it primarily colonizes rocky substrate. In this area, ROV imagery and bathymetric data also highlight the occurrence of sub-spherical rocky blocks on the seabed, often surrounded by Posidonia oceanica (Fig. 5).
Between −15 and −25 m, the Posidonia backscatter signal gradually attenuates and coralligenous bioconstructions start to be discernible. This transitional belt, which occupies about 0.37 km2, was classified as a mosaic of coralligenous and Posidonia oceanica. Visual analysis of ROV video transects, used as ground truth, indicates that in this zone, bioconstructions, mainly belonging to the bank morphotype, develop on a hard substrate that marks the widespread break in slope throughout the study area.
Below −25 m, Posidonia is no longer detected and the predominant benthic habitat is represented by coralligenous sensu stricto. These bioconstructions, often associated with fine to medium sediment and maerl, predominantly belong to the discrete relief morphotype and tend to align subparallel to the shoreline.
4.2 Extraction of coralligenous build-ups
The model extracted 12 384 polygons, but only 9211 positive morphologies were finally related to coralligenous build-ups considering the hillshade values and validation from ROV video transects collected within the study area (Fig. 7a). This means that about 25 % of the polygons extracted using the TPI were found to be artifacts after the reclassification and the polygonization of resulting raster. According to Marchese et al. (2020), the artifacts may be due to (i) occurrence of Posidonia oceanica (Innangi et al., 2015) (Fig. 8a), (ii) bad roll correction (Fig. 8c), and/or (iii) artifact concentrations on DTM boundaries (Fig. 8e). While artifacts of types (ii) and (iii) can be reduced by performing more accurate MBES surveys (i.e., larger coverage, greater overlapping, and narrower swath width), those related to Posidonia oceanica represent real morphological features that cannot be removed by improving survey quality.
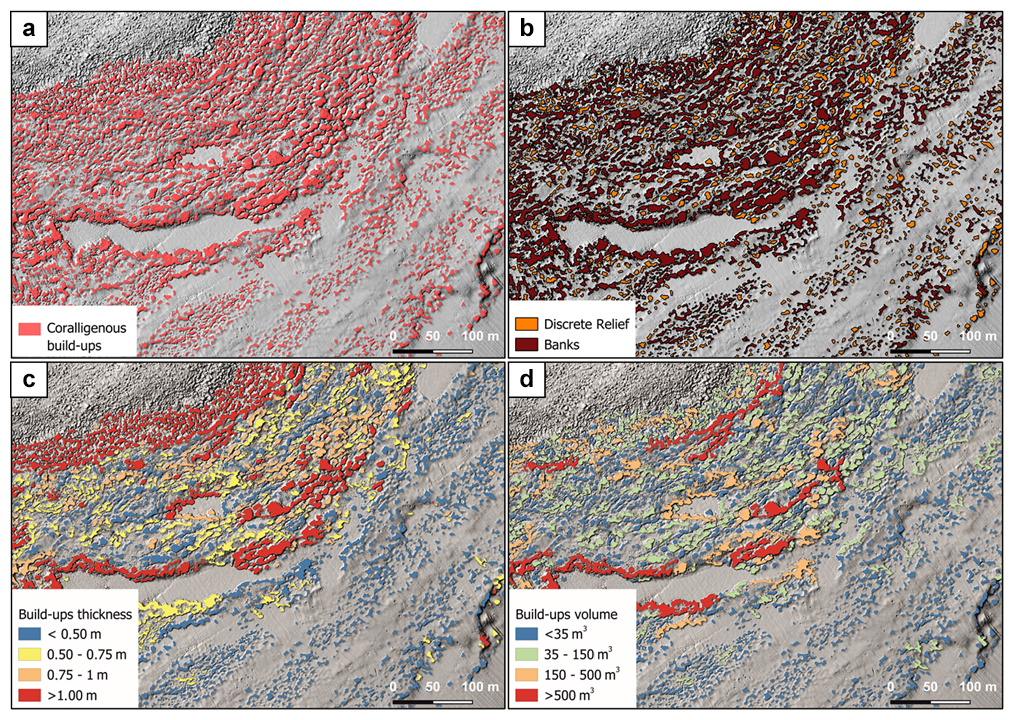
Figure 7(a) Result of build-up extraction using TPI. (b) Differentiation of coralligenous build-ups into discrete relief and banks based on the SI value. (c) Estimation of build-up thickness. (d) Calculation of the volume for each coralligenous polygon.
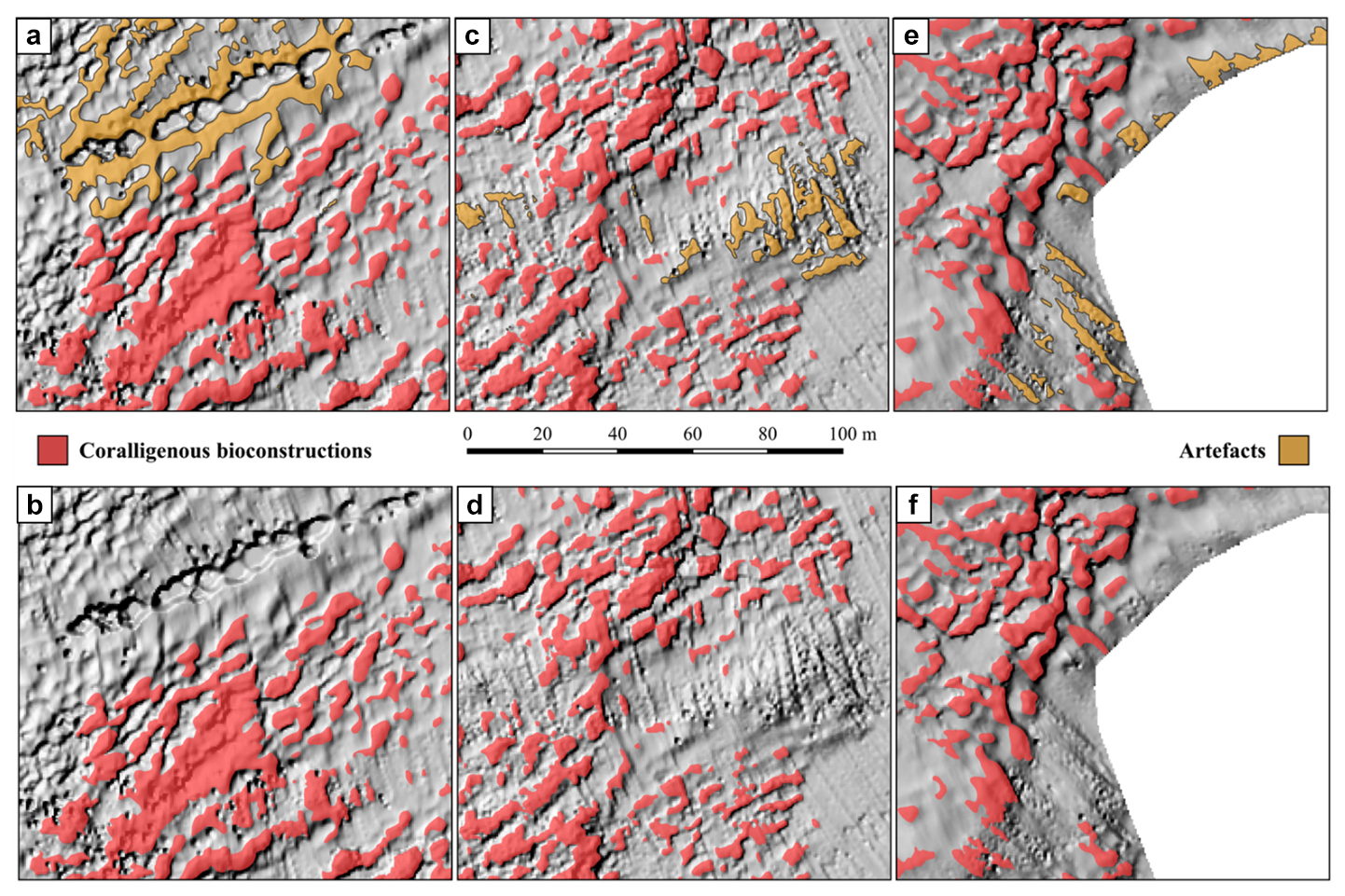
Figure 8Examples of artifacts identified during polygon extraction and their manual removal. (a) False positive caused by the presence of Posidonia oceanica and (b) the same area after removal, (c) artifact due to bad roll correction and (d) the corrected version, and (e) artifacts at the boundary of the DTM and (f) the cleaned result.
The identification of artifacts was based on specific patterns inconsistent with expected coralligenous morphologies, and their removal was carried out manually as part of the data cleaning process (Fig. 8b, d, and f). The time required for the cleaning phase strongly depends on the quality of the survey execution, the geomorphological and ecological complexity of the study area, and the experience of the operator performing the cleaning. These factors can significantly influence the extent and efficiency of manual artifact removal.
Regarding the distinction between coralligenous bioconstructions and Posidonia oceanica in the mosaic area, the separation was primarily based on the characteristics of the backscatter signal. Specifically, as discussed previously, Posidonia is associated with a moderate, speckled acoustic texture, while coralligenous bioconstructions exhibit a more complex and spatially structured acoustic signature. These interpretations were supported by ROV video transects, which help to validate the differentiation.
4.3 Shape index, thickness, surface, and volume of coralligenous build-ups
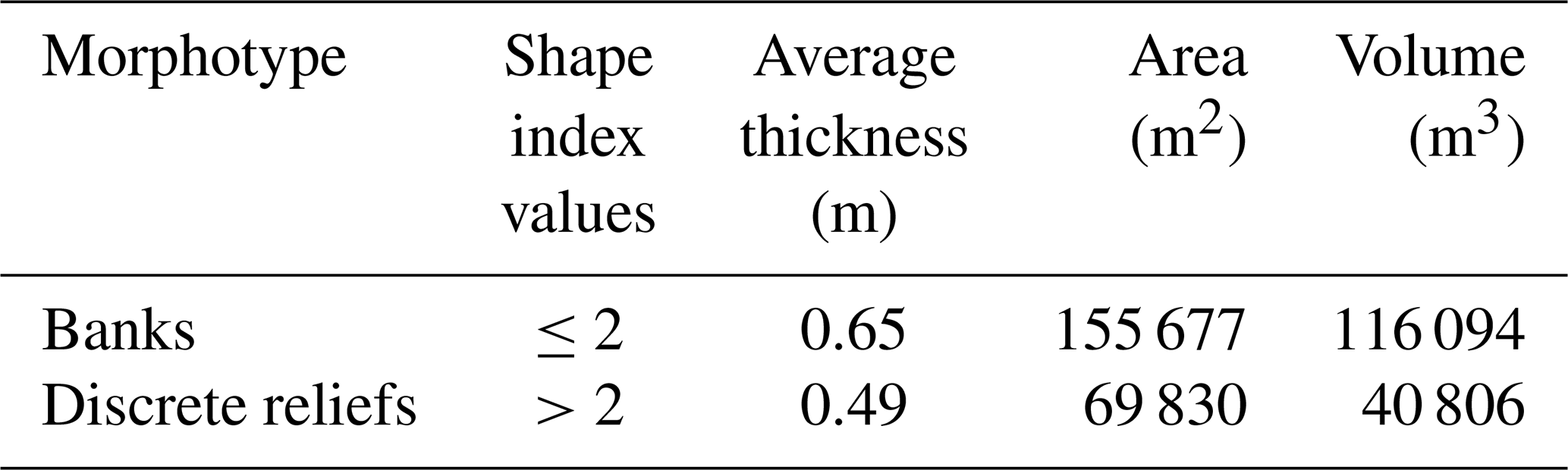
Shape index (SI) values allowed distinguishing between banks (tabular bank sensu Bracchi et al., 2016; SI ≤ 2) and discrete reliefs (discrete reliefs and hybrid banks sensu Bracchi et al., 2016; SI > 2) (Fig. 7b). Following this approach, it was possible to identify 7001 polygons belonging to the morphotype of the banks and 2210 classified as discrete reliefs. As shown in Table 1, banks have a greater average thickness (Fig. 7c) compared to discrete reliefs (0.65 m vs. 0.49 m, respectively) and cover an area of 155 677 m2, which represents about 5.2 % of the seabed in the study area. In contrast, discrete reliefs cover only 2.6 % of the seafloor, with a surface area of 69 830 m2. The volume (Fig. 7d) occupied by discrete reliefs (40 806 m3) is also significantly lower than that of the banks (116 094 m3). These data are consistent with the fact that discrete reliefs are characterized by smaller extent and thickness compared to the banks.
Table 1Classification of coralligenous polygons, based on SI values, and results in terms of area and volume.
Acoustic techniques, such as high-resolution swath bathymetry sounding (including backscatter), side scan sonar detection, and acoustic profiling, are optimal tools for quickly recognizing and identifying the extension of benthic habitats on the seabed and mapping their distribution without mechanical collection of samples, which would damage this delicate ecosystem (Bracchi et al., 2017).
Traditionally, the segmentation of MBES datasets has been performed manually, despite the fact that the process might be inaccurate and subjective (Cutter et al., 2003; Bishop et al., 2012). Initial attempts at automation employed object-oriented methods using object-based image analysis (OBIA) or considered a comprehensive set of remote data to accurately characterize seabed landforms for documenting the extension of benthic habitat (e.g., Lucieer and Lamarche, 2011; Ismail et al., 2015; Janowski et al., 2018; Fakiris and Papatheodorou, 2012). More recently, the growing availability of high-resolution MBES data has encouraged the application of deep learning approaches, particularly convolutional neural networks (CNNs) and fully convolutional neural networks (FCNNs), which produce pixel-wise classifications in order to create semantically segmented maps. These methods have proven effective in identifying geomorphological features such as bedrock outcrops, pockmarks, and submarine dune and ridges, offering high accuracy and repeatability (Arosio et al., 2023; Garone et al., 2023). Additionally, 3D CNNs have been applied to automated denoising of MBES data, enhancing the efficiency of bathymetric data workflows (e.g., Stephens et al., 2020).
Nonetheless, a universally accepted and standardized methodology for geomorphological classification of the seafloor is still lacking. Indeed, existing approaches remain highly case-specific, depending on the study area, data quality, and research objective. Moreover, relatively limited attention has been devoted to the morphological characterization of coralligenous bioconstructions, despite their ecological relevance. Indeed, only a few studies have attempted to map these complex biogenic structures in detail. Bracchi et al. (2017) proposed a categorization of coralligenous morphotypes on subhorizontal substrate based on integrated acoustic data and ground truthing, defining new morphological classes such as tabular banks, hybrid banks, and discrete reliefs across the Apulian shelf. Subsequently, Marchese et al. (2020) proposed a protocol that combines acoustic datasets and geomorphometric analysis, performed using ArcGISTM, in order to define the 2D and 3D complexity of coralligenous build-ups and to quantify how much carbonate is deposited. More recently, Varzi et al. (2023) produced a morpho-bathymetric map for the continental shelf offshore of Marzamemi (Sicily, Italy) that contained a quantitative description for the distribution and extent of coralligenous reefs.
The approach proposed in this work, based on the workflow shown in Fig. 2, represents the first attempt to define the benthic habitat in the Isola Capo Rizzuto Marine Protected Area and to quantify the extent and morphometric characteristics of coralligenous bioconstructions present therein using exclusively open-source software during post-processing phases.
5.1 Spatial distribution of benthic habitats and seafloor morphology
The benthic habitat distribution identified in the study area exhibits a clear spatial zonation, which appears to be influenced by both substrate characteristics and geomorphological features. In the shallowest sector (above −15 m depth), Posidonia oceanica represents the prevalent benthic habitat. In the intermediate depth range (down to approximately −25 m depth), a mosaic of Posidonia and coralligenous bioconstructions develops, indicating a transitional zone where environmental conditions allow the coexistence of seagrass and algal reefs.
Comparison between the morphological characteristics of the seabed with the alignment and elevation of the emerged marine terraces highlights the presence of a flat, laterally continuous submerged surface, as typically observed in relict marine terraces (e.g., Savini et al., 2021; Lebrec et al., 2022). This subhorizontal platform is bounded seaward by a break in slope, located at approximately −15 m depth, interpreted as the outer margin of the terrace. Based on this evidence, the submerged surface can be correlated with the fifth-order terrace exposed near Le Castella, characterized by a gently seaward-inclined surface and a morphological step interpreted as a paleocliff (Bracchi et al., 2016). The different orientation of the submerged scarp in the study area (NE–SW), compared to the emerged paleocliff associated with the Le Castella marine terrace (NW–SE to E–W), may be reasonably attributed to local coastal curvature and/or tectonic influences. The submersion of this portion of the fifth-order terrace in the study area would be justified by the possible presence of a tectonic feature with extensional kinematics located approximately along the coastline, which shows a distinctly straight alignment with a N–S orientation. However, further investigations are needed to confirm this hypothesis.
The inner portion of the submerged surface is characterized by the presence of sub-spherical blocks, often colonized by Posidonia oceanica, which possibly result from gravitational processes affecting the adjacent fourth-order marine terrace located upslope. This interpretation is supported by their rounded morphology, typically associated with detachment and downslope transport, and by the presence of scarps in the emerged portion of the study area, which could indicate past gravitational instability.
The outer portion and the edge of the submerged platform (down to approximately −25 m) host several coralligenous build-ups, predominantly belonging to the bank morphotype. Similar spatial arrangements have been observed in submerged terraces of southeastern Sicily (Varzi et al., 2023) and on wave-cut ravinement surfaces associated with fossil marine terraces, such as the mid-Pleistocene Cutro terrace (Nalin et al., 2006) and the emerged fifth-order terrace of Le Castella (Bracchi et al., 2016).
In the deeper sector of the study area (below −25 m depth), Posidonia is no longer present and the benthic assemblages are composed of coralligenous sensu stricto associated with fine to medium sediments and maerl. These bioconstructions mainly belong to the discrete relief morphotype and tend to follow a subparallel orientation relative to the shoreline, a distribution pattern that appears to be associated with relatively pronounced seafloor structures (as revealed by ROV video transects). This spatial configuration suggests that environmental or geomorphological factors may influence the development and positioning of build-ups. Particularly, two hypotheses are proposed to explain this pattern: (i) the influence of bottom currents and internal waves, which may promote the alignment of coralligenous bioconstructions, as observed in mesophotic carbonate systems of the Maltese shelf by Bialik et al. (2024), and (ii) an overprint of the build-ups onto inherited seabed morphologies, shaped by sea level fluctuation and regional uplift during the Quaternary glacial–interglacial cycles, as documented on submerged terraces offshore Marzamemi (SE Sicily) by Varzi et al. (2023). However, further investigations, including in situ hydrodynamic and sediment transport measurements, are necessary to validate these hypotheses.
5.2 TPI-based feature extraction
Coralligenous build-ups were treated as distinct features in both two- and three-dimensional spaces, with the aim of using geomorphometric parameters for their extraction from the seafloor. Variability of coralligenous morphotypes (Bracchi et al., 2017) poses several challenges for their automated extraction from the DTM. Since build-ups rise from the surrounding seafloor, their detection could be performed by slope analysis. However, while slope proves effective for accurately segmenting isolated small-scale features (Savini et al., 2014; Bargain et al., 2017), it struggles to incorporate the inner areas of banks into the segmentation process. The high 3D complexity in these areas makes it challenging to create a continuous polygon. On the other hand, geomorphometric parameters like the rugosity index (i.e., TRI; Riley et al., 1999; Micallef et al., 2012) are more successful in defining the overall distribution of bank morphotypes, but they fail to provide an accurate estimation of the size of discrete reliefs. Therefore, as noted by Marchese et al. (2020), TPI offers a good compromise for detecting coralligenous morphotypes. Indeed, it assesses the relative topographic position of a central point by calculating the difference between its elevation and the average elevation within a predefined neighborhood. In this work, the input parameters for the calculation of the TPI have been refined in order to minimize the artifacts during the extraction process. Specifically, the choice of a threshold value of 0.2 (lower than 0.3 used by Marchese et al., 2020), combined with higher values of power and bandwidth compared to the default ones, has allowed for a 15 % reduction in the artifact percentage compared to Marchese et al. (2020). These adjustments have therefore significantly reduced the manual review time, improving the automatization of the extraction process.
The threshold value adopted for the TPI analysis was defined through a trial-and-error procedure, as described in the methodological section. In particular, threshold values lower than 0.2 increased the morphological adherence of the extracted features to seabed forms, but at the cost of a higher number of false positives (especially in areas covered by Posidonia oceanica, where slight topographic variations were incorrectly interpreted as relevant morphotypes). Conversely, threshold values higher than 0.2 reduced the occurrence of artifacts but led to the omission of low-relief structures, thus compromising the completeness of mapping. In this work, a threshold value of 0.2 proved to be an effective compromise, ensuring a satisfactory balance between the accuracy of morphotype extraction and the minimization of false positive. This configuration allowed for the preservation of relevant coralligenous bioconstructions, including low-relief build-ups, while significantly limiting the occurrence of artifacts.
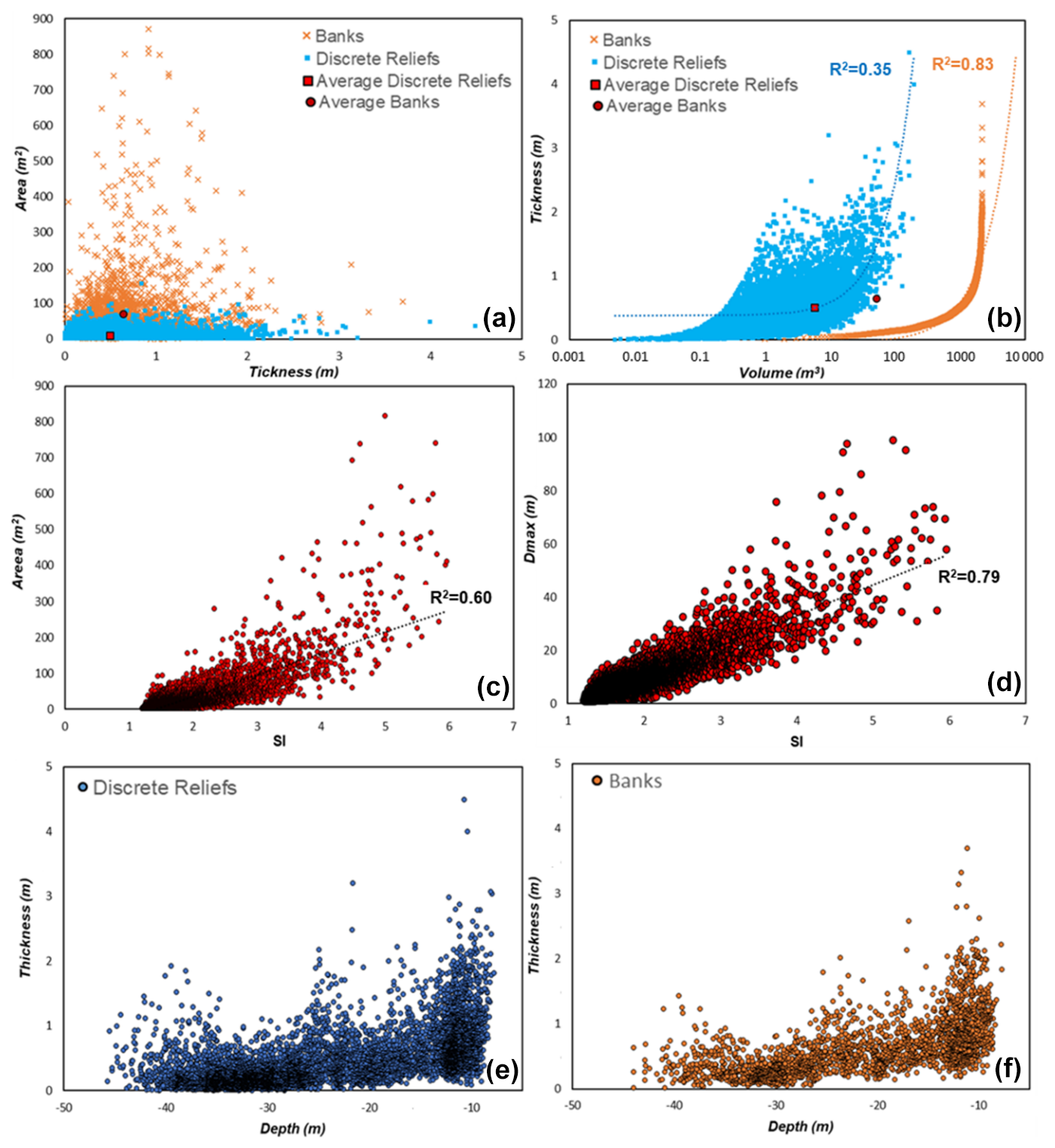
Figure 9Scatterplot representing relationships between area and thickness (a), thickness and volume (b), area and shape index (c), maximum diameter and shape index (d), thickness and depth for banks (e), and discrete relief (f). These quantitative geometric data were extracted by the benthic habitat mapping model proposed in this work. SI: shape index; Dmax: maximum diameter.
The proposed approach, although developed only for a specific coastal area, can be transferred to other regions, provided that adequate calibration is performed. The effectiveness of TPI-based extraction depends on several factors, and no universally applicable threshold value exists, as it must be adapted to the resolution and quality of bathymetric data, as well as to the site-specific geomorphological and geobiological variability. To date, no standardized procedure is available for determining the optimal threshold; however, its selection can be refined through iterative testing supported by ground-truth validation. Once the appropriate input parameter for TPI calculation (e.g., power, bandwidth, minimum and maximum radius) and a suitable threshold value are identified, the method allows for the extraction of morphologically distinct features, provided these are sufficiently expressed relative to the surrounding seafloor.
5.3 Morphological development of coralligenous build-ups
The quantitative morphometric data (i.e., surface, thickness, volume, maximum diameter, and shape indices), extracted from the benthic habitat mapping model proposed in this work, are plotted in the scatterplots of Fig. 9, providing new insights into the spatial distribution, morphotype variability, and growth pattern of the coralligenous build-ups across the study area.
Most polygons, representing aggregates of different coralligenous build-ups, are characterized by areas smaller than 200 m2 and less than 1 m thick (Fig. 9a). However, discrete reliefs and banks display some differences in their distribution: discrete reliefs tend to cluster in the lower part of the graph (smaller areas and lower thickness), whereas banks with similar thickness generally exhibit larger areas on average.
The volume of the build-ups is strongly dependent on thickness, suggesting that vertical growth plays a key role in the formation of these structures (Fig. 9b). However, discrete reliefs show a more irregular distribution, with a greater dispersion of data (R2=0.35). This trend suggests that volume increase depends not only on thickness but also on a significant lateral growth component. Conversely, banks exhibit a more regular trend, with volume increasing proportionally with thickness. The strong correlation between thickness and volume (R2=0.83) aligns with a growth pattern that is almost exclusively vertical for this morphotype.
The relationships between the area and shape index (SI) of coralligenous build-ups (Fig. 9c), despite a moderate data dispersion, revealed a positive correlation (R2=0.61), suggesting that more irregularly shaped bioconstructions (typically associated with the morphotypes of banks) tend to cover larger areas. Moreover, banks also tend to have larger maximum diameter (Dmax), as suggested by an R2 value of 0.78 (Fig. 9d). However, the greater variability in area might reflect higher spatial complexity in the distribution of these structures.
The relationship between depth and thickness of coralligenous bioconstructions, divided into banks (Fig. 9f) and discrete reliefs (Fig. 9e), reveals that both morphotypes exhibit average decreasing thickness with increasing depth. However, discrete reliefs show greater thickness variability, with higher dispersion of data at depths shallower than −25 m, whereas for the banks, data distribution is more regular. The decrease in the thickness of bioconstructions with increasing depth could be attributed to various causes, including changes in hydrodynamic energy, the characteristics of the substrate on which the bioconstructions develop, or sedimentation conditions.
To date, no previous study has provided morphometric analysis of coralligenous build-ups based on quantitative extraction of 2D/3D parameters (e.g., area, thickness, volume, shape indices) from high-resolution MBES data. Therefore, a direct comparison of our results with other Mediterranean coralligenous fields is currently not possible. Nonetheless, several works have described the geomorphological variability of coralligenous morphotypes across the Mediterranean basin (e.g., Bracchi et al., 2015, 2017, 2022; Marchese et al., 2020). These studies recognize the coexistence of morphotypes such as banks and discrete reliefs, often occurring over short spatial scales and associated with different environmental conditions. The same spatial mixing of these morphotypes that may be due to small-scale variations in substrate type, hydrodynamic regime, or inherited seabed features, which locally favor distinct growth modes despite spatial proximity (Bracchi et al., 2017; Marchese et al., 2020; Varzi et al., 2023), was also observed in our study area.
A new mapping approach starting from high-resolution acoustic data acquired through MBES surveys performed offshore of Capo Bianco (Isola Capo Rizzuto Marine Protected Area) was developed and presented here. The method represents a step forward, as it builds on two integrated foundational approaches in coralligenous habitat studies: the morphotyping of coralligenous based on the shape index and their spatial and volumetric quantification.
The innovation of this work lies in the synthesis of these methodologies, which were applied and refined in a new study area. Moreover, the approach, which integrates bathymetric and backscatter data with geomorphological and geomorphometric indices, was performed using open-source software, providing a detailed workflow that can be freely reproduced and adopted by organizations involved in research, monitoring, and conservation of marine habitats.
The resulting model proved capable not only of identifying and differentiating the benthic habitats but also of providing new quantitative information regarding the spatial distribution and 2D/3D geometric characteristics of the extracted coralligenous build-ups. This innovative aspect, compared to the traditional mapping protocol, is crucial for the quantification of the structural complexity of these bioconstructions. Moreover, this approach enables monitoring of variations not only in terms of the habitat's areal extent, but also in terms of vertical development of coralligenous relative to the substrate from which build-ups form. Indeed, the quantitative geomorphometric data obtained from the mapping model of Capo Bianco seafloor were analyzed, revealing significant insights into the covered surface, volume, and thickness of build-ups, as well as the relationships among these parameters. In particular, the results highlighted that the discrete relief morphotype exhibits a much more pronounced lateral growth component compared to the banks. If confirmed through accurate geobiological characterization, these findings could provide important new insights about the tempo and mode of the inception and development of these hard-biogenic substrates, crucial for the conservation of Mediterranean biodiversity.
The datasets needed to evaluate the results and conclusions in this paper are available at http://geocube.unical.it//gmaruca/Dataset_Benthic_Habitat_Mapping.zip (Maruca et al., 2025). The raw data used in this study were acquired through MBES surveys using a pole-mounted, Norbit WBMS Basic multibeam sonar system integrated with GNSS/INS (Applanix OceanMaster). The processing of MBES bathymetric data was performed using QPS Qimera (http://geocube.unical.it//gmaruca/Dataset_Benthic_Habitat_Mapping.zip, last access: 4 September 2025; Maruca et al., 2025). Backscatter data processing was performed using QPS Fledermaus (http://geocube.unical.it//gmaruca/Dataset_Benthic_Habitat_Mapping.zip, last access: 4 September 2025; Maruca et al., 2025). Figures 1, 3, 4, 6, 7, and 8 were made with QGIS 3.34.9 Prizren software (http://geocube.unical.it//gmaruca/Dataset_Benthic_Habitat_Mapping.zip, last access: 4 September 2025; Maruca et al., 2025). Figure 9 was generated using Microsoft Excel (http://geocube.unical.it//gmaruca/Dataset_Benthic_Habitat_Mapping.zip, last access: 4 September 2025; Maruca et al., 2025). Data used to generate the figures are available upon request to the corresponding author.
Conceptualization: GM, AG. Methodology: GM, AG, GI, FM. Formal analysis and investigation: GM, MC, GI, US, FM. Writing – original draft preparation: GM, MC, GV, FP, AL, EC, RS. Writing – review and editing: RD, CA, FB, VAB, DB, AR, AG. Funding acquisition: AG, FB. Resources: RD, FB, AL, EC, AG. Supervision: AG.
The contact author has declared that none of the authors has any competing interests.
This work reflects only the authors' views and opinions; neither the Ministry for University and Research nor the European Commission can be considered responsible for them.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
The authors would like to thank the editor Damian Leonardo Arévalo-Martínez, the reviewer Peter Feldens, and one anonymous reviewer. We would also like to express our sincere gratitude to the Geobiology and Marine Laboratories of the DiBEST, University of Calabria, for their invaluable support and contribution to this work. This is Catania Paleontological Research Group contribution no. 529.
This work was funded by the Next Generation EU – “Technologies for climate change adaptation and quality of life improvement – Tech4You” project, as well as “Development of tools and applications for integrated marine communities and substrates monitoring”, PP 2.3.1 – Action 1, “Development of hardware and software systems for three-dimensional detection, sampling and mapping of underwater environments” (CUP H23C22000370006).
This paper was edited by Damian Leonardo Arévalo-Martínez and reviewed by Peter Feldens and one anonymous referee.
Abdullah, M. Z., Chuah, L. F., Zakariya, R., Syed, A., Rozaimi, C. H., Mahmud, S. M., Abdallah M. E., Bokhari, A., Muhammad, S. A., and Al-Shwaiman, H. A.: Evaluating climate change impacts on reef environments via multibeam echosounder and Acoustic Doppler Current profiler technology, Environ. Res., 252, 118858, https://doi.org/10.1016/j.envres.2024.118858, 2024.
Arosio, R., Hobley, B., Wheeler, A. J., Sacchetti, F., Conti, L. A., Furey, T., and Lim, A.: Fully convolutional neural networks applied to large-scale marine morphology mapping, Front. Mar. Sci., 10, 1228967, https://doi.org/10.3389/fmars.2023.1228867, 2023.
Ballesteros, E.: Mediterranean Coralligenous Assemblages: a synthesis of present knowledge, Annu. Rev. Oceanogr. Mar. Biol., 44, 123–195, 2006.
Bargain, A., Marchese, F., Savini, A., Taviani, M., and Fabri, M. C.: Provincia di Santa Maria di Leuca (Mar Mediterraneo): Identificazione di cumuli adatti all'insediamento di coralli in acque fredde utilizzando proxy geomorfometrici e metodi maxent. Front. Mar. Sci., 4, 338, https://doi.org/10.3389/fmars.2017.00338, 2017.
Basso, D., Bracchi, V. A., Bazzicalupo, P., Martini, M., Maspero, F., and Bavestrello, G.: Living coralligenous as geo-historical structure built by coralline algae, Front. Earth Sci., 10, 961632, https://doi.org/10.3389/feart.2022.961632, 2022.
Bazzicalupo, P., Cipriani, M., Guido, A., Bracchi, V. A., Rosso, A., and Basso, D.: Calcareous nannoplankton inside coralligenous build-ups: the case of Marzamemi (SE, Sicily), Bollettino della Società Paleontologica Italiana, 63, 89–99, https://doi.org/10.4435/BSPI.2024.09, 2024.
Bazzicalupo, P., Bracchi, V. A., Cipriani, M., Guido, A., Rosso, A., Sanfilippo, R., Maspero, F., Galli, A., de Ponti, E., and Basso, D.: A thousand-year journey of lithification: CT-analysis and radiocarbon dating of algal reef from Central Mediterranean. Mar. Geol., 489. https://doi.org/10.1016/j.margeo.2025.107637, 2025.
Belluomini, G., Gliozzi, E., Ruggieri, G., Branca, M., and Delitala, L.: First dates on the terraces of the Cortone Peninsula (Calabria, southern Italy), Ital. J. Geosci., 107, 249–254, 1988.
Bertolino, M., Calcinai, B., Cattaneo-Vietti, R., Cerrano, C., Lafratta, A., Pansini, M., Pica, D., and Bavestrello, G.: Stability of the Sponge Assemblage of Mediterranean Coralligenous Concretions Along a Millennial Time Span, Mar. Ecol., 35, 149–158. https://doi.org/10.1111/maec.12063, 2014.
Betzler, C., Brachert, T. C., Braga, J. C., and Martin, J. M.: Nearshore, temperate, carbonate depositional systems (lower Tortonian, Agua Amarga Basin, southern Spain): Implications for carbonate sequence stratigraphy, Sediment. Feol., 113, 27–53, 1977.
Bialik, O. M., Coletti, G., Berndt, C., Schmidt, M., and Micallef, A.: Controlli sulle facies carbonatiche mesofotiche e sulla distribuzione dei sedimenti nella piattaforma maltese, Mar Mediterraneo centrale. Facies 70, 16, https://doi.org/10.1007/s10347-024-00690-1, 2024.
Bishop, M. P., James, L. A., Shroder, J. F., and Walsh, S. J.: Geospatial technologies and digital geomorphological mapping: Concepts, issues and research, Geomorphology, 137, 5–26, https://doi.org/10.1016/j.geomorph.2011.06.027, 2012.
Bonardi, G., Cavazza, W., Perrone, V., and Rossi, S.: Calabria-Peloritani terrane and northern Ionian Sea, in: Anatomy of an Orogen: The Apennines and Adjacent Mediterranean Basins, edited by: Vai, G. B. and Martini, I. P., Kluwer Academic Publishers, 287–306, https://doi.org/10.1007/978-94-015-9829-3_17, 2001.
Bracchi, V. A., Nalin, R., and Basso, D.: Paleoecology and dynamics of coralline-dominated facies during a Pleistocene transgressive-regressive cycle (Capo Colonna marine terrace, Southern Italy), Palaeogeogr. Palaeocl. Palaeoecol., 414, 296–309, https://doi.org/10.1016/j.palaeo.2014.09.016, 2014.
Bracchi, V. A., Savini, A., Marchese, F., Palamara, S., Basso, D., and Corselli C.: Coralligenous habitat in the Mediterranean Sea: a geomorphological description from remote data, Ital. J. Geosci., 134, 32–40, https://doi.org/10.3301/IJG.2014.16, 2015.
Bracchi, V. A., Nalin, R., and Basso, D.: Morpho-structural heterogeneity of shallow-water coralligenous in a Pleistocene marine terrace (Le Castella, Italy), Palaeogeogr. Palaeocl. Palaeoecol., 454, 101–112, https://doi.org/10.1016/j.palaeo.2016.04.014, 2016.
Bracchi, V. A., Basso, D., Marchese, F., Corselli, C., and Savini, A.: Coralligenous morphotypes on subhorizontal substrate: A new categorization, Cont. Shelf Res., 144, 10–20, https://doi.org/10.1016/j.csr.2017.06.005, 2017.
Bracchi, V. A., Bazzicalupo, P., Fallati, L., Varzi, A. G., Savini, A., Negri, M. P., Rosso, A., Sanfilippo, R., Guido, A., Bertolino, M., Costa, G., De Ponti, E., Leonardi, R., Muzzupappa, M., and Basso, D.: The Main Builders of Mediterranean Coralligenous: 2D and 3D Quantitative Approaches for its Identification, Front. Earth Sci., 10, 910522, https://doi.org/10.3389/feart.2022.910522, 2022. Bracchi, V. A., Negri, M. P., Bazzicalupo, P., Bertolino, M., Cipriani, M., Donato, G., Guido, A., Rosso, A., Sanfilippo, R., Sciuto, F. and Basso, D.: Mollusk diversity of coralligenous build-ups in the southwestern Ionian Sea. Mediterr. Mar. Sci., https://doi.org/10.12681/mms.38878, 2025.
Brown, C. J., Sameoto, J. A., and Smith, S. J.: Multiple methods, maps, and management applications: Purpose made seafloor maps in support of ocean management, J. Sea Res., 72, 1–13, https://doi.org/10.1016/j.seares.2012.04.009, 2012.
Cavazza, W., Blenkinsop, J., De Celles, P. G., Patterson, R. T., and Reinhardt, E. G.: Stratigrafia e sedimentologia della sequenza sedimentaria oligocenica-quaternaria del bacino Calabro-Ionico, Bollettino della Società Paleontologica Italiana, 116, 51–77, 1997.
Chiocci, F. L., Budillon, F., Ceramicola, S., Gamberi, F., and Orrù, P.: Atlante dei lineamenti di pericolosità geologica dei mari italiani, CNR edizioni, RM: Risultati del progetto MaGIC, ISBN 978-88-8080-457-4, 2021.
Cipriani, M., Basso, D., Bazzicalupo, P., Bertolino, M., Bracchi, V. A., Bruno, F., Costa, G., Dominici, R., Gallo, A., Muzzupappa, M., Rosso, A., Perri, F., Sanfilippo, R., Sciuto, F., and Guido, A.: The role of non-skeletal carbonate component in Mediterranean Coralligenous: new insight from the CRESCIBLUREEF project, Rendiconti Online Societa Geologica Italiana, 59, 75–79, https://doi.org/10.3301/ROL.2023.12, 2023.
Cipriani, M., Apollaro, C., Basso, D., Bazzicalupo, P., Bertolino, M., Bracchi, V. A., Bruno, F., Costa, G., Dominici, R., Gallo, A., Muzzupappa, M., Rosso, A., Sanfilippo, S., Sciuto, F., Vespasiano, G., and Guido, A.: Origin and role of non-skeletal carbonate in coralligenous build-ups: new geobiological perspectives in biomineralization processes, Biogeosciences, 21, 49–72, https://doi.org/10.5194/bg-21-49-2024, 2024.
Conrad, O., Bechtel, B., Bock, M., Dietrich, H., Fischer, E., Gerlitz, L., Wehberg, J., Wichmann, V., and Böhner, J.: System for Automated Geoscientific Analyses (SAGA) v. 2.1.4, Geosci. Model Dev., 8, 1991–2007, https://doi.org/10.5194/gmd-8-1991-2015, 2015.
Cosentino, D., Gliozzi, E., and Salvini, F.: Brittle deformations in the Upper Pleistocene deposits of the Crotone Peninsula, Calabria, southern Italy, Tectonophysics, 163, 205–217, 1989.
Cutter, G. R., Rzhanov, Y., and Mayer, L. A.: Automated segmentation of seafloor bathymetry from multibeam echosounder data using local fourier histogram texture features, J. Exp. Mar. Biol. Ecol., 285, 355–370, https://doi.org/10.1016/S0022-0981(02)00537-3, 2003.
De Falco, G., Tonielli, R., Di Martino, G., Innangi, S., Simeone, S., and Parnum, I. M.: Relationships between multibeam backscatter, sediment grain size and Posidonia oceanica seagrass distribution, Cont. Shelf Res., 30, 1941–1950, https://doi.org/10.1016/j.csr.2010.09.006, 2010.
De Falco, G., Conforti, A., Brambilla, W., Budillon, F., Ceccherelli, G., and De Luca, M.: Coralligenous banks along the western and northern continental shelf of Sardinia Island (Mediterranean Sea), J. Maps, 18, 200–209, https://doi.org/10.1080/17445647.2021.2020179, 2022.
Deias, C., Guido, A., Sanfilippo, R., Apollaro, C., Dominici, R., Cipriani, M., Barca, D., and Vespasiano, G.: Elemental Fractionation in Sabellariidae (Polychaeta) Biocement and Comparison with Seawater Pattern: A New Environmental Proxy in a High-Biodiversity Ecosystem?, Water, 15, 1549, https://doi.org/10.3390/w15081549, 2023.
Di Geronimo, I., Di Geronimo, R., Improta, S., Rosso, A., and Sanfilippo, R.: Preliminary observation on a columnar coralline build-up from off SE Sicily, Biologia Marina Mediterranea, 8, 229–237, 2001.
Donato, G., Sanfilippo, R., Basso, D., Bazzicalupo, P., Bertolino, M., Bracchi, V. A., Cipriani, M., D'Alpa, F., Guido, A., Negri, M. P., Sciuto, F., Serio, D., and Rosso, A.: Biodiversity associated with a coralligenous build-up off Sicily (Ionian Sea), Reg. Stud. Mar. Sci., 80, 103868, https://doi.org/10.1016/j.rsma.2024.103868, 2024.
Faccenna, C., Becker, T. W., Lucente, F. P., Jolivet, L., and Rossetti, F.: History of subduction and back-arc extension in the Central Mediterranean, Geophys. J. Int., 145, 809–820, https://doi.org/10.1046/j.0956-540x.2001.01435 https://doi.org/10.1046/j.0956-540x.2001.01435.x, 2001.
Faccenna, C., Molin, P., Orecchio, B., Olivetti, V., Bellier, O., Funiciello, F., Minelli, L., Piromallo, C., and Billi, A.: Topography of the Calabria subduction zone (Southern Italy): clues for the origin of Mt. Etna, Tectonics, 30, TC1003, https://doi.org/10.1029/2010TC002694, 2011.
Fakiris, E. and Papatheodorou, G.: Quantification of regions of interest in swath sonar backscatter images using grey-level and shape geometry descriptors: The TargAn software, Mar. Geophys. Res., 33, 169–183, https://doi.org/10.1007/s11001-012-9153-5, 2012.
Ferrigno, F., Rendina, F., Sandulli, R., and Fulvio Russo, G.: Coralligenous assemblages: research status and trends of a key Mediterranean biodiversity hotspot through bibliometric analysis, Ecol. Quest., 35, 19–36, https://doi.org/10.12775/EQ.2024.002, 2024.
Foglini, F., Grande, V., Marchese, F., Bracchi, V. A., Prampolini, M., Angeletti, L., Castellan, G., Chimienti, G., Hansen, I. M., Gudmundsen, M., Meroni, A. N., Mercorella, A., Vertino, A., Badalamenti, F., Corselli, C., Erdal, I., Martorelli, E., Savini, A., and Taviani, M.: Application of Hyperspectral Imaging to Underwater Habitat Mapping, Southern Adriatic Sea, Sensors, 19, 2261, https://doi.org/10.3390/s19102261, 2019.
Fonseca, L. and Mayer, L.: Remote estimation of surficial seafloor properties through the application of angular range analysis to multibeam sonar data, Mar. Geophys. Res., 28, 119–126, https://doi.org/10.1007/s11001-007-9019-4, 2007.
Garone, R. V., Lønmo, T. I. B., Schimel, A. C. G., Diesing, M., Thorsnes, T., and Løvstakken, L.: Seabed classification of multibeam echosounder data into bedrock/non-bedrock using deep learning, Front. Earth Sci., 11, 1285368, https://doi.org/10.3389/feart.2023.1285368, 2023.
Gerovasileiou, V. and Bianchi, C. N.: Mediterranean marine caves: a synthesis of current knowledge, in: Oceanography and Marine Biology: An Annual Review, Vol. 59, edited by: Hawkins, S. J., Lemasson, A. J., Allcock, A. L., Bates, A. E., Byrne, M., Evans, A. J., Firth, L. B., Marzinelli, E. M., Russell, B. D., Smith, I. P., Swearer, S. E., and Todd, P. A., Taylor & Francis, 1–88, https://doi.org/10.1201/9781003138846-1, 2021.
Gliozzi, E.: I terrazzi marini del Pleistocene superiore della penisola di Crotone (Calabria), Geologica Romana, 26, 17–79, 1987.
Guido, A., Heindel, K., Birgel, D., Rosso, A., Mastandrea, A., Sanfilippo, R., Russo, F., and Peckmann, J.: Pendant bioconstructions cemented by microbial carbonate in submerged marine cave (Holocene, SE Sicily), Palaeogeogr. Palaeocl. Palaeoecol., 388, 166–180, https://doi.org/10.1016/j.palaeo.2013.08.007, 2013.
Guido, A., Rosso, A., Sanfilippo, R., Russo, F., and Mastandrea, A.: Frutexites from microbial/metazoan bioconstructions of recent and Pleistocene marine caves (Sicily, Italy), Palaeogeogr. Palaeocl. Palaeoecol., 453, 127–138, https://doi.org/10.1016/j.palaeo.2016.04.025, 2016.
Guido, A., Rosso, A., Sanfilippo, R., Russo, F., and Mastandrea, A.: Microbial Biomineralization in Biotic Crusts from a Pleistocene Marine Cave (NW Sicily, Italy), Geomicrobiol. J., 34, 864–872, https://doi.org/10.1080/01490451.2017.1284283, 2017a.
Guido, A., Jimenez, C., Achilleos, K., Rosso, A., Sanfilippo, R., Hadjioannou, L., Petrou, A., Russo, F., and Mastandrea, A.: Cryptic serpulid-microbialite bioconstructions in the Kakoskali submarine cave (Cyprus, Eastern Mediterranean), Facies, 63, 21, https://doi.org/10.1007/s10347-017-0502-3, 2017b.
Guido, A., Gerovasileiou, V., Russo, F., Rosso, A., Sanfilippo, R., Voultsiadou, E., and Mastandrea, A.: Composition and biostratinomy of sponge-rich biogenic crusts in submarine caves (Aegean Sea, Eastern Mediterranean), Palaeogeogr. Palaeocl. Palaeoecol., 534, 109338, https://doi.org/10.1016/j.palaeo.2019.109338, 2019a.
Guido, A., Gerovasileiou, V., Russo, F., Rosso, A., Sanfilippo, R., Voultsiadou, E., and Mastandrea, A.: Dataset of biogenic crusts from submarine caves of the Aegean Sea: An example of sponges vs microbialites competitions in cryptic environments, Data Brief, 27, 104745, https://doi.org/10.1016/j.dib.2019.104745, 2019b.
Guido, A., Rosso, A., Sanfilippo, R., Miriello, D., and Belmonte, G.: Skeletal vs microbialite geobiological role in bioconstructions of confined marine environments, Palaeogeogr. Palaeocl. Palaeoecol., 593, 110920, https://doi.org/10.1016/j.palaeo.2022.110920, 2022.
Ingrosso, G., Abbiati, M., Badalamenti, F., Bavestrello, G., Belmonte, G., Cannas, R., Benedetti Cecchi, L., Bertolino, M., Bevilacqua, S., Bianchi, C. N., Bo, M., Boscari, E., Cardone, F., Cattaneo Vietti, R., Cau, A., Cerrano, C., Chemello, R., Chimienti, G., Congiu, L., Corriero, G., Costantini, F., De Leo, F., Donnarumma, L., Falace, A., Fraschetti, S., Giangrande, A., Gravina, M. F., Guarnieri, G., Mastrototaro, F., Milazzo, M., Morri, C., Musco, L., Pezzolesi, L., Piraino, S., Prada, F., Ponti, M., Rindi, F., Russo, G. F., Sandulli, R., Villamor, A., Zane, L., and Boero, F.: Mediterranean Bioconstructions Along the Italian Coast, Adv. Mar. Biol., 79, 61–136, https://doi.org/10.1016/bs.amb.2018.05.001, 2018.
Innangi, S., Barra, M., Di Martino, G., Parnum, I. M., Tonielli, R., and Mazzola, S.: Reson SeaBat 8125 backscatter data as a tool for seabed characterization (Central Mediterranean, Southern Italy): Results from different processing approaches, Appl. Acoust., 87, 109–122, https://doi.org/10.1016/j.apacoust.2014.06.014, 2015.
Innangi, S., Ferraro, L., Innangi, M., Di Martino, G., Giordano, L., Bracchi, V. A., and Tonielli, R.: Linosa island: a unique heritage of Mediterranean biodiversity, J. Maps, 20, 2297989, https://doi.org/10.1080/17445647.2023.2297989, 2024.
Ismail, K., Huvenne, V. A. I., and Masson, D. G.: Objective automated classification technique for marine landscape mapping in submarine canyons, Mar. Geol., 362, 17–32, https://doi.org/10.1016/j.margeo.2015.01.006, 2015.
Janowski, L., Trzcinska, K., Tegowski, J., Kruss, A., Rucinska-Zjadacz, M., and Pocwiardowski, P.: Nearshore Benthic Habitat Mapping Based on Multi-Frequency, Multibeam Echosounder Data Using a Combined Object-Based Approach: A Case Study from the Rowy Site in the Southern Baltic Sea, Remote Sens., 10, 1983, https://doi.org/10.3390/rs10121983, 2018.
Jardim, V. L., Grall, J., Barros-Barreto, M. B., Bizien, A., Benoit, T., Braga, J. C., Brodie, J., Burel, T., Cabrito, A., Diaz-Pulido, G., Gagnon, P., Hall-Spencer, J. M., Helias, M., Horta, P. A., Joshi, S., Kamenos, N. A., Kolzenburg, R., Krieger, E. C., Legrand, E., Page, T. M., Peña, V., Ragazzola, F., Rasmusson, L. M., Rendina, F., Schubert, N., Silva, J., Tâmega, F. T. S., Tauran, A., and Burdett, H. L.: A Common Terminology to Unify Research and Conservation of Coralline Algae and the Habitats They Create, Aquat. Conserv., 35, e70121, https://doi.org/10.1002/aqc.70121, 2025.
Laborel, J.: Marine biogenic constructions in the Mediterranean. A review, Scientific Reports of Port-Cros National Park 13, https://doi.org/10.1111/j.1365-2117.2012.00549.x, 97–126, 1987.
Lamarche, G. and Lurton, X.: Recommendations for improved and coherent acquisition and processing of backscatter data from seafloor-mapping sonars, Mar. Geophys. Res., 39, 5–22, https://doi.org/10.1007/s11001-017-9315-6, 2018.
Lebrec, U., Riera, R., Paumard, V., Leary, M. J. O., and Lang, S. C..: Morphology and distribution of Submerged palaeoshorelines: Insights from the North West Shelf of Australia, Earth-Sci. Rev., 224, 103864, https://doi.org/10.1016/j.earscirev.2021.103864, 2022.
Lecours, V., Devillers, R., Schneider, D. C., Lucieer, V. L., Brown, C. J., and Edinger, E. N.: Spatial scale and geographic context in benthic habitat mapping: Review and future directions, Mar. Ecol.-Prog. Ser., 535, 259–284, https://doi.org/10.3354/meps11378, 2015.
Lecours, V., Dolan, M. F. J., Micallef, A., and Lucieer, V. L.: A review of marine geomorphometry, the quantitative study of the seafloor, Hydrol. Earth Syst. Sci., 20, 3207–3244, https://doi.org/10.5194/hess-20-3207-2016, 2016.
Lo Iacono, C., Savini, A., and Basso, D.: Cold-Water carbonate bioconstructions, in: Submarine geomorphology, edited by: Micallef, A., Krastel, S., and Savini, A., Springer, 425–455, ISBN 425-3-319-57851-4, https://doi.org/10.1007/978-3-319-57852-1_22, 2018.
Lucieer, V. and Lamarche, G.: Unsupervised fuzzy classification and object-based image analysis of multibeam data to map deep water substrates, Cook Straight, New Zealand, Cont. Shelf Res., 31, 1236–1247, https://doi.org/10.1016/j.csr.2011.04.016, 2011.
Lurton, X. Lamarche, G., Brown, C., Lucieer, V., Rice, G., Schimel, A., and Weber, T. (Eds): Backscatter Measurements by Seafloor-mapping Sonars, Guidelines and Recommendations, GeoHab, Zenodo [data set], https://doi.org/10.5281/zenodo.10089261, 2015.
Malinverno, A. and Ryan, W. B. F.: Extension in the Tyrrhenian Sea and shortening in the Apennines as result of arc migration driven by sinking of the lithosphere, Tectonics, 5, 227–245, https://doi.org/10.1029/TC005i002p00227, 1986.
Marchese, F., Bracchi, V. A., Lisi, G., Basso, D., Corselli, C., and Savini, S.: Assessing Fine-Scale Distribution and Volume of Mediterranean Algal Reefs through Terrain Analysis of Multibeam Bathymetric Data. A Case Study in the Southern Adriatic Continental Shelf, Water, 12, 157, https://doi.org/10.3390/w12010157, 2020.
Maruca, G., Cipriani, M., Dominici, R., Imbrogno, G., Vespasiano, G., Apollaro, C., Perri, F., Bruno, F., Lagudi, A., Severino, U., Bracchi, V. A., Basso, D., Cellini, E., Mauri, F., Rosso, A., Sanfilippo, R., and Guido, A.: Dataset Benthic Habitat Mapping, geocube [data set], http://geocube.unical.it//gmaruca/Dataset_Benthic_Habitat_Mapping.zip, 2025.
Massari, F. and Prosser, G.: Late Cenozoic tectono-stratigraphic sequences of the Crotone Basin: insights on the geodynamic history of the Calabrian arc and Tyrrhenian Sea, Basin Res., 25, 26–51, https://doi.org/10.1111/j.1365-2117.2012.00549.x, 2013.
Mauz, B. and Hassler, U.: Luminescence chronology of late Pleistocene raised beaches on Southern Italy: new data on relative sea-level changes, Mar. Geol., 170, 187–203, https://doi.org/10.1016/S0025-3227(00)00074-8, 2000.
McGarigal, K. and Marks, B. J. F.: Spatial Pattern Analysis Program for Quantifying Landscape Structure (General Technical Report), https://doi.org/10.2737/PNW-GTR-351, Washington, D.C., USA, 1995.
Micallef, A., Le Bas, T. P., Huvenne, V. A. I., Blondel, P., Hühnerbach, V., and Deidun, A.: A multi-method approach for benthic habitat mapping of shallow coastal areas with high-resolution multibeam data, Cont. Shelf Res., 39, 14–26, https://doi.org/10.1016/j.csr.2012.03.008, 2012.
Milia, A. and Torrente, M. M.: Early-stage rifting of the southern Tyrrhenian region: the Calabria-Sardinia breakup, J. Geodynam., 81, 17–29, https://doi.org/10.1016/j.jog.2014.06.001, 2014.
Minelli, L. and Faccenna, C.: Evolution of the Calabrian accretionary wedge (Central Mediterranean), Tectonics, 29, TC4004, https://doi.org/10.1029/2009TC002562, 2010.
MSFD, CE: Direttiva 2008/56/CE del Parlamento europeo e del Consiglio, del 17 giugno 2008, che istituisce un quadro per l'azione comunitaria nel campo della politica per l'ambiente marino (direttiva quadro sulla strategia per l'ambiente marino, Bruxelles), 2008.
Nalin, R. and Massari, F.: Facies and stratigraphic anatomy of a temperate carbonate sequence (Capo Colonna Terrace, late Pleistocene, Southern Italy), J. Sediment. Res., 79, 210–225, https://doi.org/10.2110/jsr.2009.027, 2009.
Nalin, R., Basso, D., and Massari, F.: Pleistocene coralline algal build-ups (coralligéne de plateau) and associated bioclastic deposits in the sedimentary cover of Cutro marine terrace (Calabria, Southern Italy), in: Cool-Water Carbonates: Depositional Systems and Palaeoenvironmental Controls, edited by: Pedley, H. M. and Carannante, G., The Geological Society of London, 11–22, https://doi.org/10.1144/GSL.SP.2006.255.01.02, 2006.
Nalin, R., Massari, F., and Zecchin, M.: Superimposed cycles of composite marine terraces: the example of Cutro Terrace (Calabria, Southern Italy), J. Sediment. Res., 77, 340–354, https://doi.org/10.2110/jsr.2007.030, 2007.
Nalin, R., Bracchi, V. A., Basso, D., and Massari, F.: Persististrombus latus (Gmelin) in the upper Pleistocene deposits of the marine terraces of the Crotone peninsula (Southern Italy), Ital. J. Geosci. 131, 95–101, https://doi.org/10.3301/IJG.2011.25, 2012.
Palmentola, G., Carobene, L., Mastronuzzi, G., and Sansò, P.: I terrazzi marini pleistocenici della Penisola di Crotone (Italia), Geografia Fisica e Dinamica Quaternaria, 13, 75–80, 1990.
Pepe, F., Sulli, A., Bertotti, G., and Cella, F.: Architecture and Neogene to Recent evolution of the western Calabrian continental margin: An upper plate perspective to the Ionian subduction system, central Mediterranean, Tectonics, 29, TC3007, https://doi.org/10.1029/2009TC002599, 2010.
Pérès, J. M.: Structure and dynamics of assemblages in the benthal, Mar. Ecol., 5, 119–185, 1982.
Pérès, J. M. and Picard, J.: Nouveau manuel de bionomie benthique de la Mer Méditerranée, Recent Travaux de la Station Marine d'Endoume, 31, 5–137, 1964.
Picone, F. and Chemello, R.: Seascape characterization of a Mediterranean vermetid reef: a structural complexity assessment, Front. Mar. Sci., 10, 1134385, https://doi.org/10.3389/fmars.2023.1134385, 2023.
Reitz, M. A. and Seeber, L.: Arc-parallel strain in a short rollback-subduction system: the structural evolution of the Crotone basin (Northeastern Calabria, Southern Italy), Tectonics, 31, TC4017, https://doi.org/10.1029/2011TC003031, 2012.
Riley, S. J., De Gloria, S. D., and Elliot, R.: A Terrain Ruggedness Index that Quantifies Topographic Heterogeneity, Int. J. Scient. Res., 5, 23–27, 1999.
Rosso, A., Donato, G., Sanfilippo, R., Serio, D., Sciuto, F., D'Alpa, F., Bracchi, V. A., Negri, M. P., and Basso, D.: The bryozoan Margaretta cereoides as a habitat-former in the Coralligenous of Marzamemi (SE Sicily, Mediterranean Sea), in: Marine Benthic Biodiversity of Eastern Mediterranean Ecosystems, edited by: Koulouri, P., Gerovasileiou, V., and Dailianis, T., J. Mar. Sci. Eng., 11, 590, https://doi.org/10.3390/jmse11030590, 2023.
Rueda, J. L., Urra, J., Aguilar, R., Angeletti, L., Bo, M., García-Ruiz, C., Gonzalez-Duarte, M. M., Lopez, E., Madurell, T., Maldonado, M., Mateo-Ramirez, A., Megina, C., Moreira, J., Moya, F., Ramalho, L. V., Rosso, A., Sitjà, C., and Taviani, M.: Cold-Water Coral Associated Fauna in the Mediterranean Sea and Adjacent Areas, in: Mediterranean Cold-Water Corals: Past, Present and Future, Coral Reefs of the World, Vol. 9, edited by: Orejas, C. and Jiménez, C., Springer International Publishing AG, part of Springer Nature, 295–333, https://doi.org/10.3390/jmse11030590, 2019.
Sanfilippo, R., Rosso, A., Mastandrea, A., Viola, A., Deias, C., and Guido, A.: Sabellaria alveolata sandcastle worm from the Mediterranean Sea: New insights on tube architecture and biocement, J. Morphol., 280, 1839–1849, https://doi.org/10.1002/jmor.21069, 2019.
Sanfilippo, R., Rosso, A., Viola, A., Guido, A., and Deias, C.: Architecture and tube structure of Sabellaria spinulosa (Leuckart, 1849): comparison with the Mediterranean S. alveolata congener, J. Morphol., 283, 1350–1358, https://doi.org/10.1002/jmor.21507, 2022.
Sanfilippo, R., Donato, G., Sciuto, F., Guido, A., Cipriani, M., Bracchi, V. A., Viola, A., Basso, D., and Rosso, A.: Serpulid diversity of coralligenous build-ups in the southwestern ionian Sea. African Soology, 60(1), https://doi.org/10.1080/15627020.2024.2440954, 2025.
Santagati, P., Guerrieri, S., Borrelli, M., and Perri, E.: Calcareous bioconstructions formation during the last interglacial (MIS 5) in the central Mediterranean: A consortium of algal, metazoan, and microbial framebuilders (Capo Colonna-Crotone Basin South Italy), Mar. Petrol.Geol., 167, 106950, https://doi.org/10.1016/j.marpetgeo.2024.106950, 2024.
Santoro, E., Mazzella, M. E., Rerranti, L., Randisi, A., Napolitano, E., Rittner, S., and Radtke, U.: Raised coastal terraces along the Ionian Sea coast of Northern Calabria, Italy, suggest space and time variability of tectonic uplift rates, Quatern. Int., 206, 78–101, https://doi.org/10.1016/j.quaint.2008.10.003, 2009.
Savini, A., Vertino, A., Marchese, F., Beuck, L., and Freiwald, A.: Mapping cold-water coral habitats at different scales within the Northern Ionian Sea (central Mediterranean): An assessment of coral coverage and associated vulnerability, PLoS ONE, 9, e87108, https://doi.org/10.1371/journal.pone.0087108, 2014.
Savini, A., Borrelli, M., Vertino, A., Mazzella, F., and Corselli, C.: Terraced Landforms Onshore and Offshore the Cilento Promontory (Southern Tyrrhenian Margin): New Insights into the Geomorphological Evolution, Water, 13, 566, https://doi.org/10.3390/w13040566, 2021.
Schlager, W.: Depositional bias and environmental change-important factors in sequence stratigraphy, Sediment. Geol., 70, 109–130, 1991.
Schlager, W.: Accommodation and supply – A dual control on stratigraphic sequences, Sediment. Geol., 86, 111–136, 1993.
Sciuto, F., Altieri, C., Basso, D., D'Alpa, F., Donato, G., Bracchi, V. A., Cipriani, M., Guido, A., Rosso, A., Sanfilippo, R., Serio, D., and Viola, A.: Preliminary data on ostracods and foraminifers living on coralligenous bioconstructions Offshore Marzamemi (Ionian Sea, Se Sicily), Revue de Micropaléontologie, 18, 100711, https://doi.org/10.1016/j.revmic.2023.100711, 2023.
Severino, U., Lagudi, A., Barbieri, L., Scarfone, L., and Bruno, F.: A SLAM-Based Solution to Support ROV Pilots in Underwater Photogrammetric Survey, in:International Conference of the Italian Association of Design Methods and Tools for Industrial Engineering, Springer Nature Switzerland, Cham, 443–450, https://doi.org/10.1007/978-3-031-58094-9_49, 2023.
SNPA: Schede metodologiche utilizzate nei programmi di monitoraggio del secondo ciclo della direttiva strategia marina (D.M. 2 febbraio 2021), ISBN 978-88-448-1236-2, https://www.puntosicuro.it/_resources/files/SNPA_SchedeMetodologiche_SM_REV_compressed.pdf (last access: 5 September 2025), 2024.
Stephens, D., Smith, A., Redfern, T., Talbot, A., Lessnoff, A., and Dempsey, K.: Using three dimensional convolutional neural networks for denoising echosounder point cloud data, Appl. Comput. Geosci., 5, 100016, https://doi.org/10.1016/j.acags.2019.100016, 2020.
UNEP-MAP-RAC/SPA: Piano d'azione per la conservazione del coralligeno e di altre bioconcrezioni calcaree nel Mar Mediterraneo. Tunisi: Rac/Spa, 1–21, 2008.
UNEP-MAP-RAC/SPA: Piano d'azione per la conservazione del coralligeno e di altre bioconcrezioni calcaree nel Mar Mediterraneo. Atene, Grecia: Rac/Spa, 1–20, 2017.
Varzi, G. A., Fallati, L., Savini, A., Bracchi, V. A., Bazzicalupo, P., Rosso, A., Sanfilippo, R., Bertolino, M., Muzzupappa, M., and Basso, D.: Geomorphology of coralligenous reefs offshore southeastern Sicily (Ionian Sea), J. Maps, 19, 2161963, https://doi.org/10.1080/17445647.2022.2161963, 2023.
Vosselman, G.: Slope based filtering of laser altimetry data, Vol. XXXIII, IAPRS, Amsterdam, https://www.researchgate.net/profile/George-Vosselman/publication/228719860_Slope_based_filtering_of_laser_altimetry_data/links/00463524c7061a1220000000/Slope-based-filtering-of-laser-altimetry-data.pdf (last access: 5 September 2025), 2000.
Westaway, R.: Quaternary uplift of Southern Italy, J. Geophys. Res., 98, 21741–21772, https://doi.org/10.1029/93JB01566, 1993.
Westaway, R. and Bridgland, D.: Late Cenozoic uplift of Southern Italy deduced from fluvial and marine sediments: coupling between surface processes and lower-crustal flow, Quatern. Int., 175, 86–124, https://doi.org/10.1016/j.quaint.2006.11.015, 2007.
Wilson, J. P. and Gallant, J. C.: Analisi del terreno: principi e applicazion. (Eds.) John Wiley & Sons, Inc., pp. 479, ISBN 0-471-32188-5, 2000.
Zecchin, M. and Caffau, M.: Key features of mixed carbonate-siliciclastic shallow-marine systems: the case of Capo Colonna terrace (southern Italy), Ital. J. Geosci., 130, 370–379, https://doi.org/10.3301/IJG.2011.12, 2011.
Zecchin, M., Nalin, R., and Roda, C.: Raised Pleistocene marine terraces of the Crotone peninsula (Calabria, southern Italy): facies analysis and organization of their deposits, Sediment. Geol., 172, 165–185, https://doi.org/10.1016/j.sedgeo.2004.08.003, 2004.
Zecchin, M., Caffau, M., Civile, D., and Roda, C.: Facies and cycle architecture of a Pleistocene marine terrace (Crotone, southern Italy): a sedimentary response to late Quaternary, high-frequency glacio-eustatic changes, Sediment. Geol., 216, 138–157, https://doi.org/10.1016/j.sedgeo.2009.03.004, 2009.
Zecchin, M., Caffau, M., Civile, D., and Roda, C.: Anatomy of a late Pleistocene clinoformal sedimentary body (Le Castella, Calabria, southern Italy): A case of prograding spit system? Sediment. Geol., 223, 291–309, https://doi.org/10.1016/j.sedgeo.2009.11.015, 2010.
Zecchin, M., Caffau, M., Civile, D., Critelli, S., Di Stefano, A., Maniscalco, R., Muto, F., Sturiale, G., and Roda, C.: The Plio-Pleistocene evolution of the Crotone Basin (Southern Italy): interplay between sedimentation, tectonics and eustasy in the frame of Calabrian arc migration, Earth-Sci. Rev., 115, 273–303, https://doi.org/10.1016/j.earscirev.2012.10.005, 2012.
Zevenbergen, L. W. and Thorne, C. R.: Quantitative analysis of land surface topography, Earth Surf. Proc. Land., 12, 47–56, https://doi.org/10.1002/esp.3290120107, 1987.