the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 29 Aug 2025
| 29 Aug 2025
Cross-canyon variability in zooplankton backscattering strength in a river-influenced upwelling area
Macarena Díaz-Astudillo
Manuel I. Castillo
Pedro A. Figueroa
Leonardo R. Castro
Ramiro Riquelme-Bugueño
Iván Pérez-Santos
Oscar Pizarro
Gonzalo S. Saldías
Zooplankton are a key component of food webs in upwelling systems. Their distribution is influenced not only by mesoscale and climate dynamics but also by topography and local currents. Submarine canyons that incise the continental shelf can act as conduits, transporting deep, nutrient-rich waters to shallower regions and promoting coastal biological productivity. Consequently, these canyons facilitate the advection and accumulation of zooplankton. We aimed to describe the spatio-temporal variability in zooplankton distribution (using net samples and acoustic data) and their association with local currents in a long, narrow submarine canyon located in the highly productive continental shelf off central Chile. The backscattering strength (Sv), a proxy for zooplankton biomass, was highly variable on both diurnal and spatial scales. Higher Sv and abundances were found at night, following the classic diel vertical migration pattern. Zooplankton were not uniformly distributed within the canyon. In the surface and mid-depth layers, the canyon walls accumulated more zooplankton than the center, particularly during the nighttime. Within the canyon, the currents were asymmetrical and frequently reversed direction. When the positive along-canyon current was stronger on the northern slope, Sv was also higher on that wall. This pattern was especially evident in the section closer to the canyon head. We show that the Biobio Canyon is a highly dynamic environment where oceanographic conditions can rapidly shift. Our findings suggest a feasible mechanism for zooplankton retention driven mainly by along-canyon flow asymmetry and vertical migrations.
- Article
(9051 KB) - Full-text XML
-
Supplement
(3520 KB) - BibTeX
- EndNote




In the Humboldt Current System (HCS), wind-driven upwelling is the primary mechanism responsible for the high levels of primary productivity that support abundant zooplankton communities (Brink, 1983; Escribano et al., 2012; Medellín-Mora et al., 2016), even at the mesoscale or larger spatial scales along the coast (Landaeta and Castro, 2002; Yannicelli et al., 2006a; Riquelme-Bugueño et al., 2012; Díaz-Astudillo et al., 2022). Other relevant mesoscale and regional drivers of zooplankton abundance and distribution in this ecosystem are mesoscale eddies and fronts (Morales et al., 2007, 2010; Pavez et al., 2010; Riquelme‐Bugueño et al., 2015), changes in water mass distribution (Aronés et al., 2009), and remote low-frequency oscillations (Díaz-Astudillo et al., 2024). Although the influence of these processes on zooplankton dynamics is relatively well understood, sub-mesoscale mechanisms structuring zooplankton distribution, such as the interaction of coastal currents with topography (Prairie et al., 2012), have been less thoroughly investigated.
Abrupt changes in topography (e.g., seamounts, headlands, valleys, and submarine canyons) can lead to the retention or transport of zooplankton and micronekton through various physical mechanisms associated with flows interacting with topographic features (Genin, 2004). Most submarine canyons are V-shaped topographic features that interrupt the continuity of the continental shelf and/or the slope in continental margins worldwide (Harris and Whiteway, 2011). They modify the geostrophic flows that typically follow isobaths. Flows over submarine canyons are dominated by advective and ageostrophic forces, particularly over the head of the canyon (Saldías and Allen, 2020), leading to the generation of baroclinic tides, internal waves, and horizontal and vertical flows that enable the exchange of surface and deep waters through mixing and advection processes (Allen and Durrieu De Madron, 2009). Generally, the presence of submarine canyons in a continental margin significantly increases the transport of subsurface water from the slope to the inner shelf, promoting water exchanges along the cross-shore axis on a relatively small spatial scale near the canyon (Allen and Durrieu De Madron, 2009; Connolly and Hickey, 2014).
Usually, negative along-slope flows (i.e., equatorward in eastern boundaries) generate upwelling in the downstream wall of the canyon, while positive flows cause downwelling (Allen and Durrieu De Madron, 2009). In central Chile, episodes of extreme upwelling have been observed at the head of the Biobio Canyon, decoupled from the wind forcing. These episodes occurred with northward flows over the head of the canyon and with negative sea level anomalies nearshore, which was coherent with the passage of coastal trapped waves (Sobarzo et al., 2016). These low-frequency waves intensify the topographic upwelling that occurs in submarine canyons (Sobarzo et al., 2016; Saldías et al., 2021). This and other studies prove that topographically induced upwelling and downwelling respond to both wind forcing and sea level anomalies (Wang et al., 2022).
Through the upwelling of dense deep water into shallower depths, some canyons can provide nutrient inputs comparable to those provided by local wind-driven upwelling (Connolly and Hickey, 2014). If this deep, nutrient-rich water reaches the photic layer for a period long enough to stimulate primary productivity, canyons can become local hotspots of biological productivity and pelagic and benthic diversity (Genin, 2004; Fernandez-Arcaya et al., 2017; Santora et al., 2018). In addition to enhanced upwelling, increased vorticity generates surface recirculation patterns and asymmetric currents, potentially leading to the formation of cyclonic eddies near the canyon rim (Connolly and Hickey, 2014). These eddies can concentrate particles or organisms, an aggregation mechanism that has been observed in a relatively shallow submarine canyon on the western Antarctic Peninsula, where krill accumulate near the canyon head (Hudson et al., 2022b).
Particle transport through canyon-mediated currents can result in the concentration of potential prey in shallower waters, thus supporting trophic interactions and high predator aggregations (Genin, 2004). Thus, canyons are thought to be areas where accumulation of zooplankton is high and their advection is low (Vindeirinho, 1998). Therefore, submarine canyons usually serve as foraging sites for several pelagic predators, such as whales (Schoenherr, 1991; Croll et al., 2005; Moors-Murphy, 2014; Salgado Kent et al., 2021; Amano et al., 2023; Buchan et al., 2023), penguins (Clarke et al., 2006; Santora and Reiss, 2011; Schofield et al., 2013; Hudson et al., 2022b), and fish (De Leo et al., 2012; Saunders et al., 2021).
The study of submarine canyons along the eastern South Pacific margin has primarily focused on geological and physical processes, with limited attention being paid to their biological impacts (Silva and Araújo, 2021). The Biobio Canyon (BbC) is a long canyon located in the upwelling-influenced continental shelf off central Chile (Sobarzo et al., 2016; Vergara et al., 2024). The area surrounding the BbC provides important ecosystem services (Soto et al., 2022) and hosts abundant aggregations of zooplankton (Yannicelli et al., 2006b; Landaeta et al., 2008) and zooplankton predators such as whales (Cisterna-Concha et al., 2023). However, the mechanisms driving enhanced biological productivity in this canyon remain largely unknown. Thus, this study aims to (1) describe intra-diurnal variations in zooplankton abundance and backscattering strength over the BbC following an upwelling event and (2) examine canyon-driven physical processes influencing zooplankton variability.
2.1 Study area and field cruise
The BbC is a river-influenced, shelf-incising submarine canyon located on the continental shelf off central Chile (Fig. 1). The canyon originates near the mouth of the Biobio River and extends ∼ 40 km west–east in a zigzag pattern until reaching the shelf break. Beyond this point, it shifts to a northwestern orientation to connect with the submarine trench and the abyssal plain, with a total length of 134 km (Rodrigo, 2010). Its width ranges from 3 to 9 km, and its depth ranges from 20 to ∼ 1200 m.
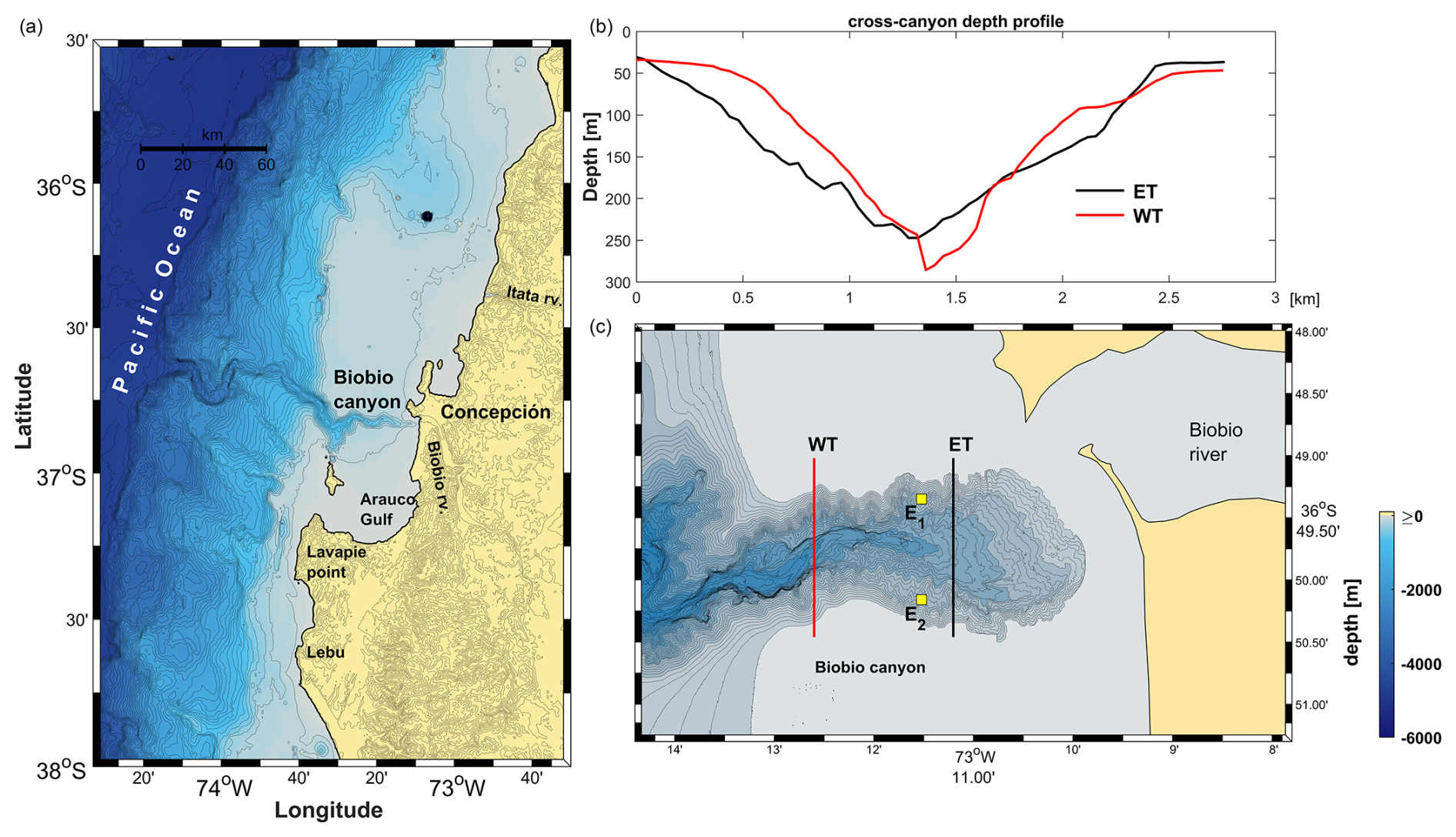
Figure 1(a) Location of the Biobio Canyon on the continental shelf and slope off central Chile. (b) Bathymetric profiles of the western transect (WT) and eastern transect (ET) are shown in red and black, respectively. (c) Zoomed-in view of the area around the canyon head, showing the positions of the WT and ET, as well as the locations of stations E1 and E2, where zooplankton samples were collected.
In general, hydrographic conditions over the shelf north of the canyon are mainly controlled by the seasonal heat flux cycle and the seasonal variability of the Biobio and Itata river discharges (Sobarzo et al., 2007). The Biobio River outflow, being the largest freshwater source in central Chile (Rojas et al., 2023), directly influences the hydrographic structure over the canyon head (Vergara et al., 2024), and its plume can influence a large extension of the Gulf of Arauco (Saldías et al., 2016; Vergara et al., 2024). Its annual cycle is mainly controlled by precipitation, resulting in higher discharges during the winter months (Saldías et al., 2012). Consequently, the river plume and riverine nutrient export are higher during the rainy season (June to September) (Masotti et al., 2018). In the region, winds are mainly driven by the Pacific anticyclone, with a seasonality marked by southwestern (SW) winds during austral spring–summer and northeastern (NE) winds during autumn–winter (Sobarzo et al., 2007; Ancapichún and Garcés-Vargas, 2015).
From 27 to 28 July 2023, we conducted a 26 h experiment over the BbC, during which we collected data on horizontal currents, acoustic backscatter, and hydrographic structure on board the RV Kay Kay II. Two cross-canyon transects (eastern and western transects; Fig. 1b, c), separated by 2.1 km, were sampled eight times each during the 26 h cycle.
2.2 Hydrographic conditions
Temperature, salinity, and chlorophyll-a (chl-a) profiles were obtained with a rapid-response towed CTD (conductivity, temperature, and depth) (Teledyne Valeport RapidPro CTD) equipped with a fluorescence sensor. The CTD was deployed during each pass along the WT and the ET (Fig. 1), completing 14 transects for the entire sampling period (Table S1 in the Supplement). Measurements extended from the surface to a depth of 100 m in the canyon or to near-bottom depths outside the canyon, with all sensors recording at a frequency of 4 Hz. The transects were conducted at intervals of no more than 2 h and were interpolated using the Barnes objective analysis scheme (Barnes, 1994).
2.3 Winds, tides, and river discharge
Hourly wind reanalysis data were obtained from the ERA5 product (https://cds.climate.copernicus.eu/, last access: 1 September 2024), the fifth-generation global climate and weather dataset from the ECMWF (Hersbach et al., 2020), at a spatial resolution of 0.25° × 0.25° (∼ 31 × 31 km). Wind components (u and v, at 10 m height) were extracted from the grid point nearest to the study site. Tide data for Concepción Bay, recorded at 1 min intervals, were downloaded from the Sea Level Monitoring Facility (https://www.ioc-sealevelmonitoring.org/list.php, last access: 1 September 2024), and provided by the Chilean Navy Hydrographic and Oceanographic Service (SHOA). Daily discharge data for the Biobio River, near its mouth, were obtained from the Dirección General de Aguas (DGA; https://snia.mop.gob.cl/dgasat/pages/dgasat_main/dgasat_main.htm, last access: 1 September 2024) to quantify freshwater input into the region.
2.4 Currents
Velocity and acoustic backscatter profiles were collected along the ET and the WT using a downward-looking 153 kHz Quartermaster Teledyne RDI acoustic Doppler current profiler (ADCP), mounted on a stainless steel arm with transducers about 1 m below the surface. Real-time measurements were acquired in bottom-track mode at a maximum vessel speed of 2 knots (2.5 m3 s−1). The ADCP was configured to collect data in 120 vertical cells (3 m bin size) at a 1 s ping rate, with a sampling time interval of 4 s. The first bin was centered at 4.31 m from the ADCP, with a profiling range of up to 350 m. Currents were recorded as zonal (u; east–west) and meridional (v; north–south) components.
Quality control of the current profiles involved removing erroneous data from each circuit, using a standard goodness-of-fit criterion greater than 50 %, to exclude flow speeds exceeding 10 m s−1, and ensuring velocity errors below 0.008 m s−1. Additionally, only profiles for which the difference between the ADCP bottom-track speed and the GPS-derived speed was less than 0.30 m s−1 were employed (e.g., Lwiza et al., 1991; Cáceres et al., 2006; Castillo et al., 2012). Directional correction was also applied since the ADCP's magnetic compass can be influenced by the vessel's magnetic field and by local magnetic anomalies (Joyce, 1989; Pollard and Read, 1989; Trump and Marmorino, 1997). Residual currents were estimated using least-squares harmonic fitting for the principal tidal components K1 (23.93 h) and M2 (12.42 h) following Lwiza et al. (1991). The results of the harmonic analysis and the residual circulation revealed that the tidal currents were weak, with amplitudes of < 0.05 m s−1.
2.5 Zooplankton backscattering strength
To investigate high-resolution variations in zooplankton abundance over the BbC, we converted the ADCP echo intensity into mean volume backscattering strength (MVBS), denoted as Sv (dB re 1 m−1). MVBS is widely used to assess zooplankton distribution and behavior, offering high-resolution data collected passively and concurrently with current measurements (Fielding et al., 2004; Dwinovantyo et al., 2019; Cisewski et al., 2021). Sv was calculated for each depth cell using the sonar equation originally proposed by Deines (1999) and modified by Mullison (2017):
where C is a sonar configuration scaling factor (−161.01 dB) that is specific to the RDI Workhorse ADCP; Tx is the temperature at the transducer (°C); L is the transmit pulse length (2.85 m); PDBW is the 10log 10 of the output power (15.72 W), α is the depth-variable sound absorption coefficient (dB m−1); E is the recorded automatic gain control (AGC or “echo counts”); and Er is the echo reference, determined as the minimum AGC recorded for each beam in the absence of any acoustic signal (40 counts). Kc is a beam-specific sensitivity coefficient used to convert the raw echo data into dB, and it was calculated following Bozzano et al. (2014) as follows:
Finally, R is the slant range in relation to the sample bin (m), which uses depth as a correction (Lee et al., 2004), and it is expressed as follows:
where B is the blanking distance (3.23 m), d is the depth cell size (3 m), n is the depth cell number of the particular scattering layer being measured, θ is the beam angle (20°), C− is the average sound speed from the transducer to the depth cell (1453 m s−1), and CI is the nominal sound speed used by the instrument (1454 m s−1). According to the ADCP's manufacturer manual, the maximum range of acceptable data (Rmax) is defined by
where H is the distance between the instrument and the bottom. To minimize potential seafloor interference, all data below Rmax were excluded. A minimum beam correlation threshold of 25 % was applied to ensure data quality. Bins where the Sv of a single beam exceeded the mean Sv of the other three beams by more than 5 dB were discarded, a procedure that enhances consistency among beams and helps eliminate signals from large scatterers (Jiang et al., 2007). Subsequently, data from the four beams were averaged, and a median filter was applied to smooth the signal. Finally, all Sv values exceeding −40 (dB m−1) were removed to further reduce the influence of large scatterers (e.g., fish with swim bladders) on the total volume backscattering strength.
After analyzing the vertical distribution of acoustic scatterers, three distinct layers were identified based on their average Sv and diurnal behavior: a surface layer extending from the surface to 25 m depth, a mid-depth layer from 25 to 100 m, and a deep layer from 100 m to the seafloor. The delineated layers were subsequently used to compare geometric-mean Sv values and horizontal-flow dynamics across different transects and slope regions. The geometric-mean allows us to compare relative changes in Sv while considering the fact that the MVBS is a logarithmic measure of backscattering cross-section per unit volume.
2.6 Zooplankton sampling
A total of eight stratified zooplankton samples were collected at stations E1 and E2 (Fig. 1c) during both the daytime and the nighttime. To avoid interfering with the continuous hydrographic and acoustic sampling conducted between 20:00 on 27 July and 22:00 on 28 July (local time is used throughout the paper), the zooplankton sampling was performed before and after the experiment (see details in Table S2). A Tucker Trawl net with a 1 m2 mouth area and a 300 µm mesh size, equipped with a General Oceanics flowmeter, was deployed to a depth of 100 m and then towed obliquely to the surface, obtaining two stratified samples from 100–50 m and 50–0 m. Samples were fixed with 5 % buffered formaldehyde for subsequent taxonomic analyses. In the laboratory, zooplankton groups were identified and quantified, with abundances standardized to individuals per 1000 m3.
3.1 Oceanographic conditions
An event of strong upwelling-favorable winds occurred prior to sampling (Fig. S1 in the Supplement). The water column was highly stratified, primarily due to freshwater input from the Biobio River. The upper 20 m were characterized by lower salinity and temperature, along with elevated chl-a concentrations (Figs. 2 and S2 to S5). In this surface layer, the eastern transect (ET), located closer to the river mouth, displayed higher chlorophyll-a levels and lower salinity and temperature than the western transect (WT) (Fig. 2a, b). Below 20 m, temperature and salinity profiles indicated the presence of Equatorial Subsurface Water (ESSW), a water mass typical of the region that intrudes onto the shelf during upwelling conditions (Sobarzo et al., 2007). At these depths, both transects exhibited similar hydrographic structures, with the most notable changes occurring around the 1025.6 kg m3 isopycnal. The WT had a slightly higher proportion of measurements within the 33.4 to 34 salinity range, which corresponded to the highest chlorophyll-a values observed at that depth (Fig. 2c, d).
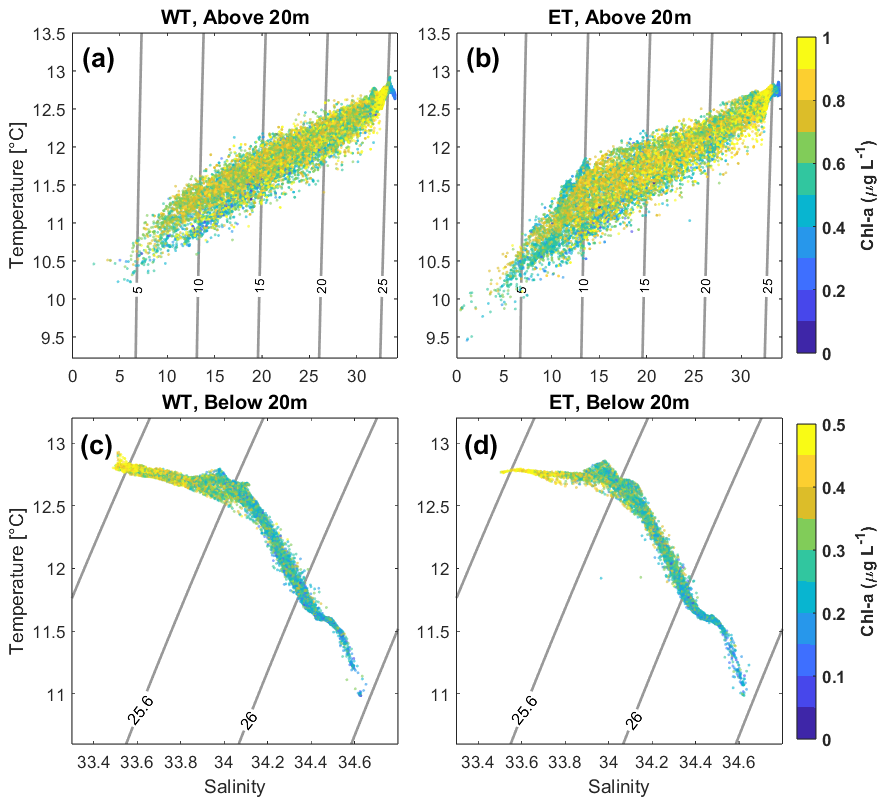
Figure 2Temperature–salinity plots for all sections along the WT (a, c) and ET (b, d). Panels (a) and (b) show the upper 20 m of the water column, while (c) and (d) represent the deeper layers. Colors indicate chlorophyll-a concentrations.
The pycnocline, defined by the 1025.6 kg m3 isopycnal, exhibited temporal and spatial variability along the cross-canyon axis (Fig. 3). Generally, it was shallower in the WT, ranging between 15 and 35 m depth, while in the ET, it varied from 12 to 30 m. In the WT, the pycnocline was approximately 10 m deeper on the southern slope (SS) of the canyon. Conversely, in the ET, it maintained a similar depth on both slopes, though it was slightly shallower on the northern slope (NS) during the initial 18 h of observation. In both transects, the pycnocline deepened over time, reaching its maximum depth by the end of the study period.
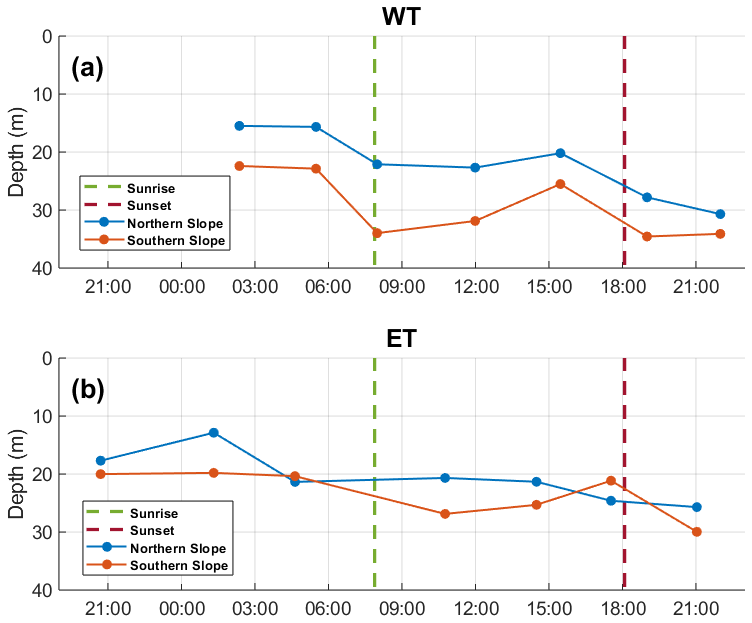
Figure 3Evolution of pycnocline depth over time in the (a) WT and (b) ET. The blue line represents the NS, and the red line represents the SS.
3.2 Horizontal flows over the canyon
The mean flow patterns in both transects are shown in Fig. 4. In both sections, the zonal (u) component exhibited a mean offshore (i.e., negative) flow from the surface down to approximately 50 m. Between 50 and ∼ 200 m, the mean flow in the WT became positive and centered within the canyon (Fig. 4a), while, in the ET, it was also positive but inclined toward the NS (Fig. 4b). Below 200 m, the flow turned negative again. However, flow variability increased considerably, as indicated by the large standard deviations in both transects (Fig. 4c, d). The meridional (v) component showed a positive (i.e., northward) surface flow in both transects, with slightly higher velocities in the WT (Fig. 4e, f), suggesting the influence of wind-driven forcing. Below 50 m, the meridional flow reversed and remained highly variable down to the bottom (Fig. 3g, h). Notably, the increased standard deviations (> 0.1 m s−1) in both the cross- and along-shore components indicate enhanced flow variability near the canyon floor.
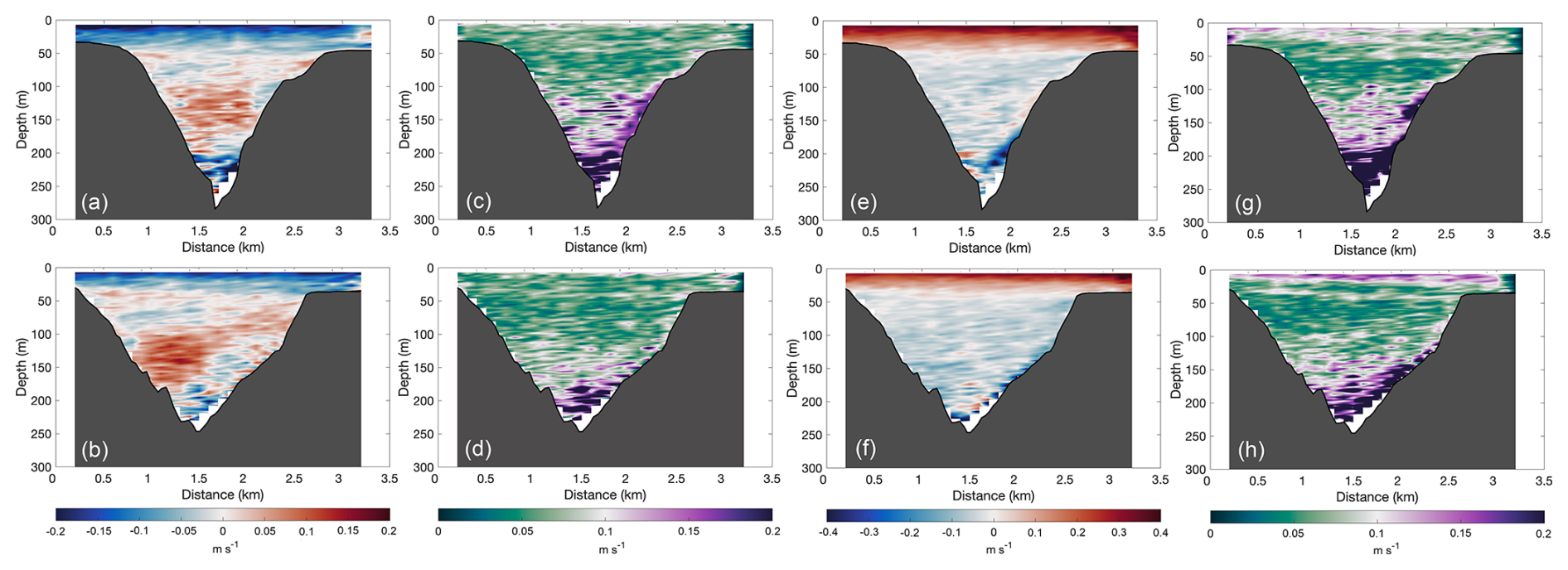
Figure 4Mean cross-shore velocities in the (a) WT and (b) ET, with their respective standard deviations shown in (c) and (d), and the mean along-shore velocities in the (e) WT and (f) ET, with their respective standard deviations shown in (g) and (h). Positive cross-shore velocities indicate onshore flow, while positive alongshore velocities indicate equatorward flow.
Spearman rank correlations between the currents on the NS and SS revealed differences in terms of the flow variability between the ET and the WT, as well as between the cross- and along-shore flow. In the ET, no significant correlations were found in the zonal velocities between the northern and southern slopes. In contrast, only the deep layer of the WT exhibited a positive correlation between the cross-shore flow between the two slopes (Fig. 5a–c). On the other hand, the along-shore velocities between the NS and SS were highly correlated across all three layers of the ET. In the surface layer, this suggests that the northward flow associated with the river plume equally affected both sides of the canyon. In the WT, a significant correlation in terms of v velocities was observed only at mid-depths (Fig. 5). These differences in the flow over the slopes suggest that the cross-shore flow is more variable inside the canyon compared to the along-shore flow. Consequently, we further investigated the variability of the cross-shore (i.e., along-canyon) component.
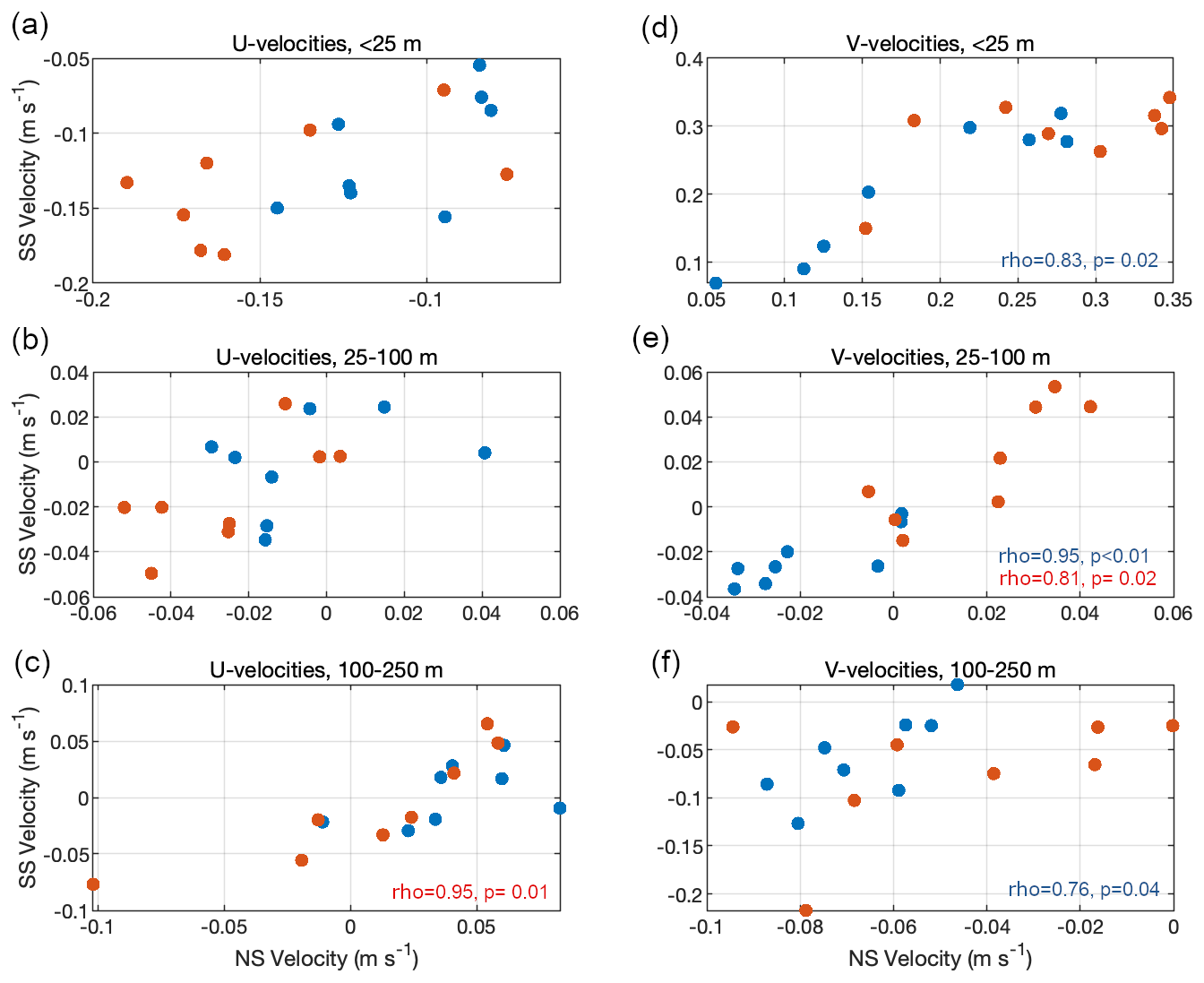
Figure 5Correlations between the mean flow along the northern slope (NS) and southern slope (SS) for both the WT (red dots) and the ET (blue dots), shown by layer. When significant, the correlation coefficient and p value are indicated.
We identified the section of the canyon where the bottom depth ranged between 60 and 120 m and computed the time–depth evolution of the cross-shore velocity at each slope. In both the NS and SS of the WT, the first 40 m of the water column exhibited negative (offshore) velocities throughout the study period. Between 40 and 120 m, the velocities were predominantly positive (onshore) along the NS, except during the first 3 h and at the end of the experiment, when the flow reversed and became strongly negative (Fig. 6a). In contrast, velocities in the 40–120 m layer of the SS remained variable, alternating between positive and negative velocities (Fig. 6b). Consequently, the velocity difference between the two slopes also fluctuated over time, indicating a stronger flow on the SS at the end of the experiment (Fig. 6c). The ET also exhibited predominantly negative velocities in the upper 40 m at both the NS and SS. Below this layer, the NS displayed mainly positive velocities throughout the experiment (Fig. 6d), whereas the SS exhibited predominantly negative velocities (Fig. 6e). The velocity difference between the slopes was variable in the upper 40 m but consistently positive in the deeper section, indicating stronger (and onshore) along-canyon currents along the NS (Fig. 6f).
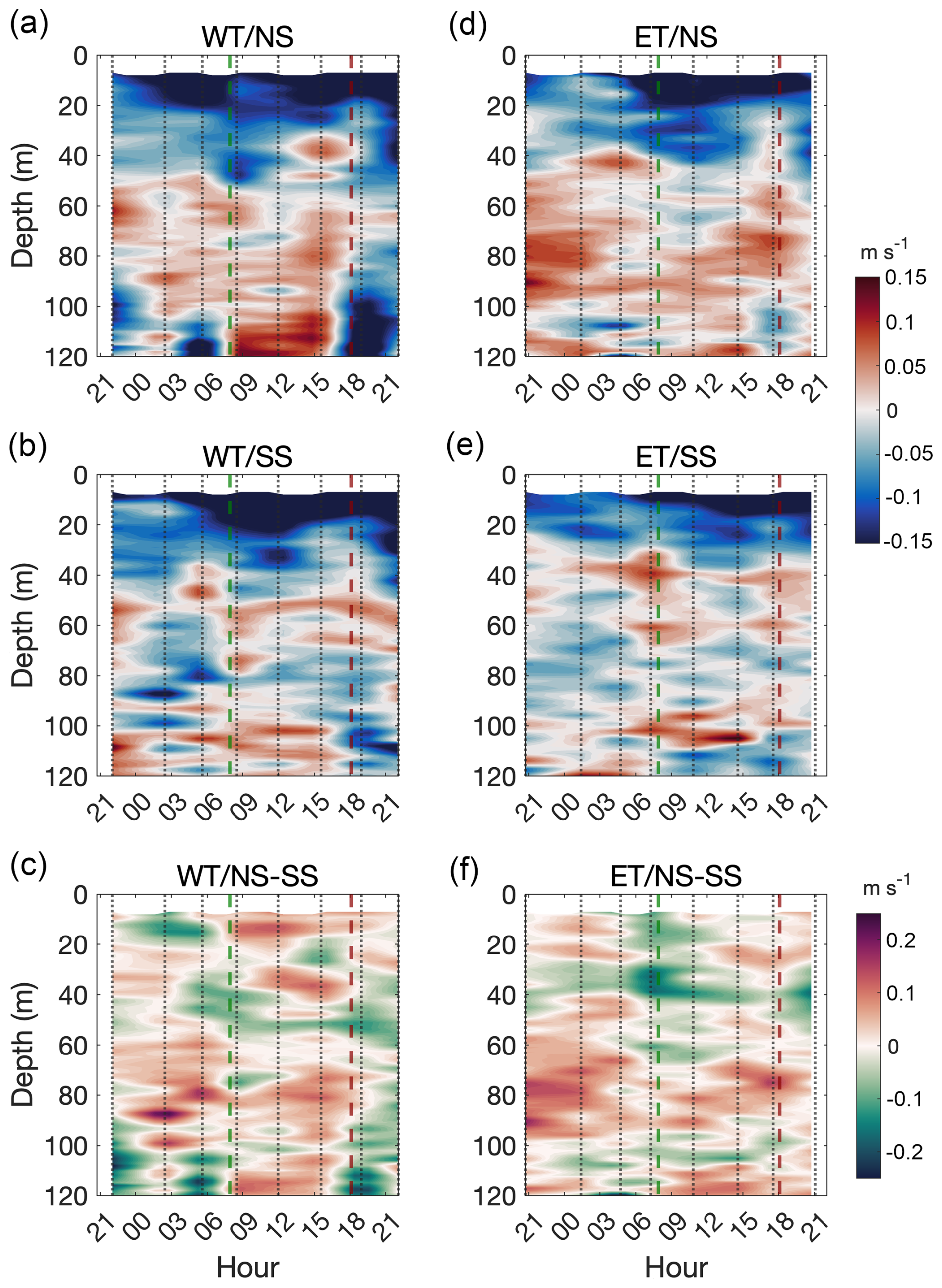
Figure 6Mean cross-shore currents by depth and time in a section of the canyon walls where bottom depth ranged from 60 to 120 m. Currents over the northern slope (NS, upper panels) and southern slope (SS, middle panels) are shown for both the WT (a, b) and the ET (d, e). The difference in current magnitude between the NS and SS is shown in (c) (WT) and (f) (ET). Vertical dotted grey lines indicate the time of each transect; the dashed green line marks the sunrise, and the dashed burgundy line marks the sunset.
3.3 Zooplankton spatial and temporal variability
Mean volume backscattering strength (Sv) over the canyon exhibited pronounced temporal, vertical, and spatial variability, both between transects and between the NS and SS. Some of the patterns observed in Sv corresponded with zooplankton abundance estimates from net tows. The most evident feature in the Sv sections was the consistent increase in Sv during nighttime hours in both the WT and the ET (Figs. 7 and S6). Zooplankton samples collected near the ET also showed higher nighttime abundances across most taxonomic groups (Fig. 7). Differences in Sv between the NS and SS closely mirrored cross-canyon differences in zooplankton abundance from net samples. For instance, toward the end of the experiment, standardized zooplankton abundances were higher on the SS (Fig. 7f), corresponding with significantly elevated Sv values on the same slope (Fig. 7d). These matching patterns support the reliability of acoustic measurements as a proxy for zooplankton distribution in the BbC.
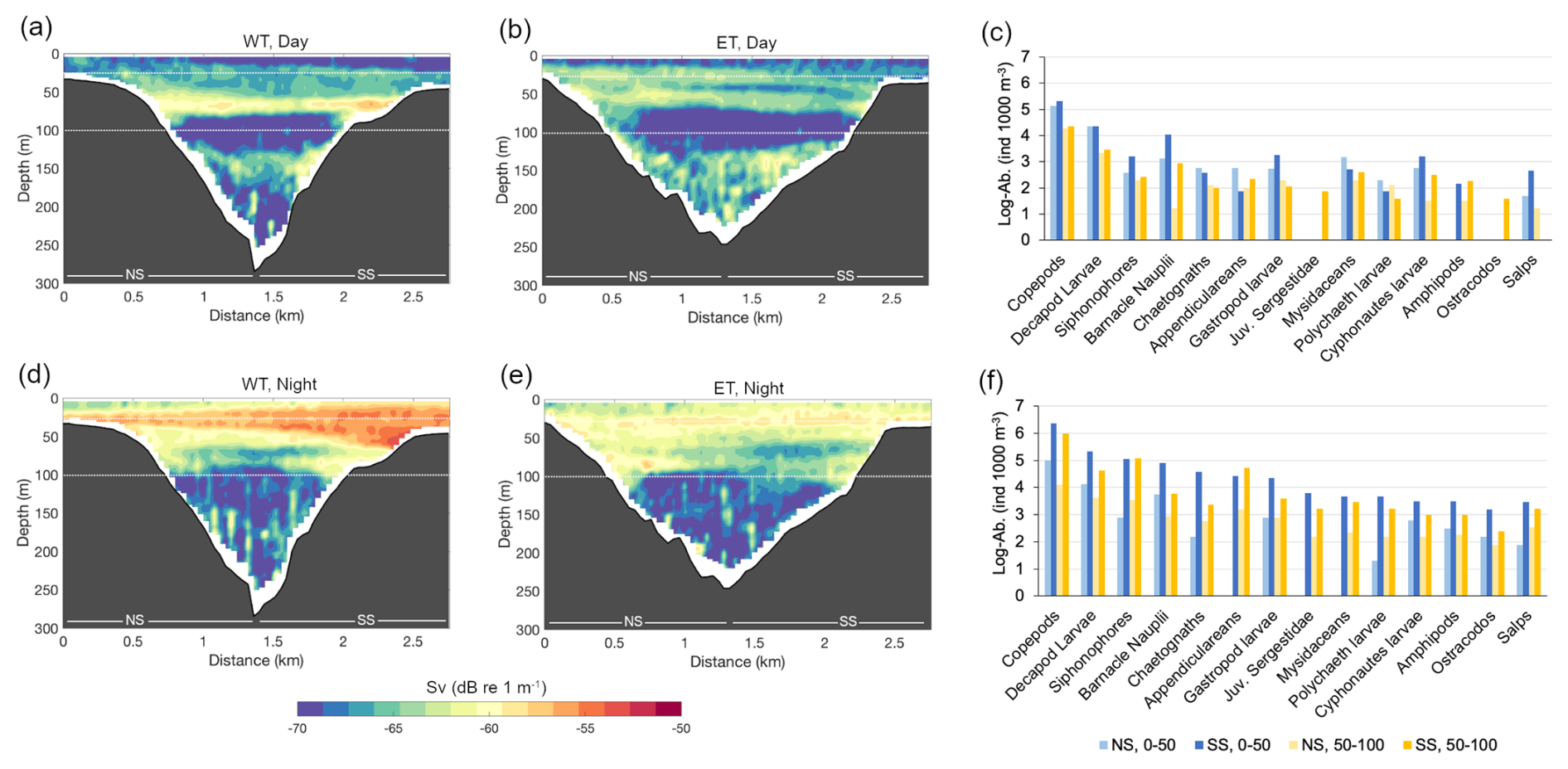
Figure 7Selected Sv sections along the western transect (WT) and eastern transect (ET). First diurnal section along the WT (a) and ET (b). Last night section along the WT (d) and ET (e). Standardized zooplankton abundances by group (from stations E1 and E2 near the ET) for samples collected during the day prior to the experiment (daytime, c) and after the end of the experiment (nighttime, f). The labels NS and SS in panels (a), (b), (d), and (e) indicate the locations of the northern and southern slopes, respectively. Horizontal dotted lines indicate the boundaries of the three layers used for layer analyses.
Diurnal and slope-related differences in Sv varied not only over time but also with depth. The surface layer exhibited pronounced intra-diurnal fluctuations, with consistently higher Sv values during the nighttime. In the WT, the difference between the lowest daytime and the highest nighttime Sv reached approximately 12 dB re m−1, whereas, in the ET, this difference increased to ∼ 18 dB re m−1 (Fig. 8a, b). In the ET, the mid-depth layer displayed the highest mean Sv values, while, in the WT, mean Sv in the mid-depth layer was comparable to that of the surface layer. In both transects, the U-shaped profiles of mean Sv values revealed an influence of canyon topography on zooplankton distribution, with higher abundances being observed over the slopes than in the central canyon axis (Fig. 8c, d). In the mid-depth layer, the diurnal signal in Sv was still evident, particularly over the canyon walls, though this was less pronounced than in the surface layer. In contrast, the deep layer showed minimal temporal variations in Sv. The clear diurnal differences observed in the upper and mid-depth layers were largely absent at depth, where Sv remained relatively constant throughout the day (Fig. 8e, f). Nevertheless, the topographic effect of the canyon was still apparent in the deep layer, with higher Sv values near the canyon walls compared to at its center.
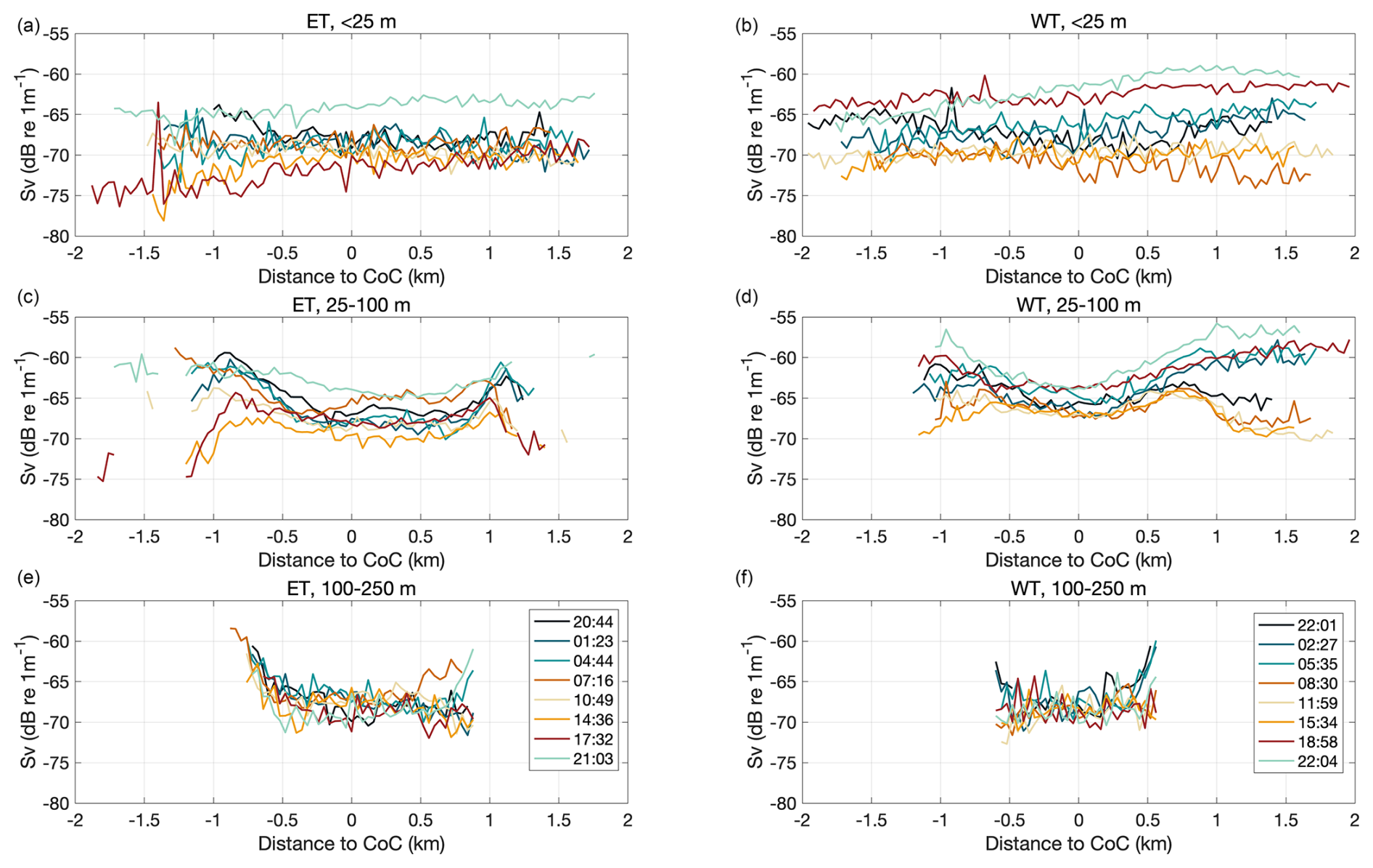
Figure 8Mean Sv in the surface (a, b), mid-depth (c, d), and deep (e, f) layers along the WT and ET. Each of the eight transects along the western (a, c, e) and eastern (b, d, f) transects is represented by a different color. Distance is referenced to the center of the canyon (CoC), defined as its deepest point, to facilitate the comparison between the NS and SS. Negative (positive) distances correspond to the northern (southern) slope.
To compare Sv between the NS and SS, we applied the same methodology previously used for u-velocity analysis. Additionally, we calculated the Sv difference between the NS and SS and associated it with horizontal current velocities in each depth layer. Distinct differences in the vertical distribution of zooplankton were observed between the slopes in both transects. In the WT, the NS consistently exhibited elevated Sv values between 20 and 80 m throughout the study period (Fig. 9a). On the SS, a ∼ 20 m thick high-Sv layer was present, which deepened during daylight hours and became shallower and thicker (up to ∼ 100 m thick) during the night (Fig. 9b). Backscatter intensity was generally higher on the SS, resulting in predominantly negative Sv differences between the NS and SS throughout the deployment (Fig. 9c).
In the ET, Sv values exceeding 63 dB re m−1 were observed across much of the NS water column during the first 12 h (Fig. 9d). Later in the day, Sv values declined in the upper 50 m, followed by a new increase near 20:00. On the SS, elevated Sv values were confined to the 20–50 m layer during the initial 12 h of the study, but this layer deepened and weakened around 09:00. (Fig. 9e). The Sv difference between the NS and SS of the ET reached ∼ 10 dB re m−1 in the 50–120 m layer at the beginning of the study, coinciding with pronounced differences in along-canyon current velocities – specifically, stronger flows were observed on the NS of the ET. Although this difference decreased over time, the NS consistently exhibited higher Sv within this depth range compared to the SS (Fig. 9f).
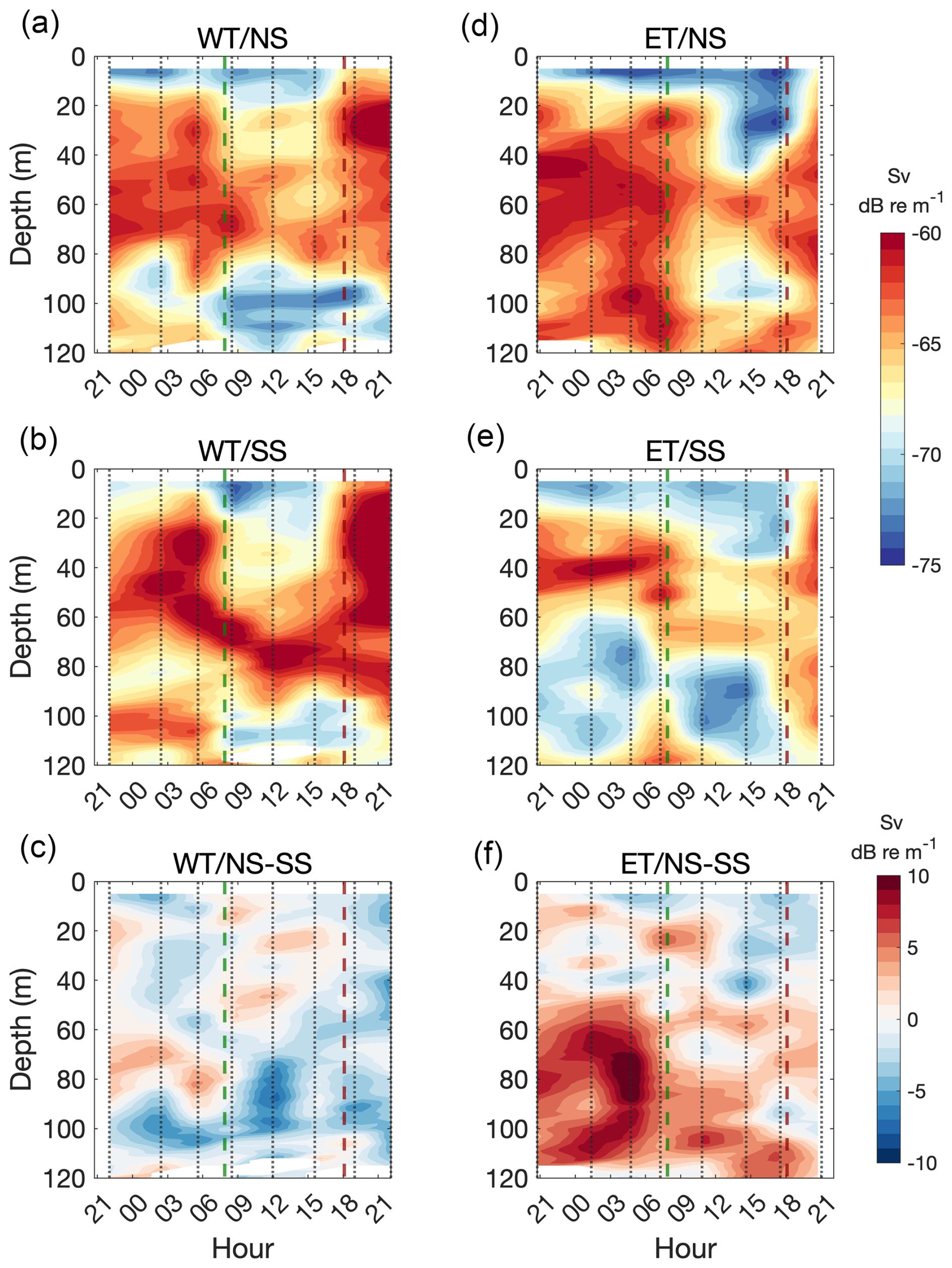
Figure 9Mean Sv currents by depth and time in a section of the canyon walls where bottom depth ranged from 60 to 120 m. Sv values over the northern slope (NS, upper panels) and southern slope (SS, middle panels) are shown for both the WT (a, b) and the ET (d, e). The difference in Sv between the NS and SS is shown in (c) (WT) and (f) (ET). Vertical dotted gray lines indicate the time of each transect, the dashed green line marks the sunrise, and the dashed burgundy line marks the sunset.
The “canyon hypothesis” suggests three main mechanisms by which submarine canyons promote local biological productivity. The first involves the fertilization of surface and subsurface layers via the advection of deeper, nutrient-rich waters to the surface (i.e., “topographic upwelling”). For this mechanism to support biological productivity, the upwelling must persist long enough to allow phytoplankton and zooplankton populations to grow and accumulate. The second mechanism involves the generation of a subsurface eddy that induces isopycnal doming, thereby enhancing upward transport and/or promoting the retention of particles. The third mechanism involves the physical retention and aggregation of organisms due to the interaction of canyon topography with alongshore currents and diel vertical migration (DVM) of planktonic organisms (Hudson et al., 2022a, b; Genin, 2004). Globally, direct observational evidence supporting these mechanisms remains limited. In our study, the short duration of observations precludes confirmation of the first mechanism, and we found no clear evidence of eddy formation within the canyon. Nonetheless, we present strong indications that the BbC plays a significant role in modulating zooplankton distribution on diurnal timescales, likely through a variation of the third mechanism.
4.1 The interaction of diel vertical migrations with abrupt topography
The intra-diurnal changes in Sv within the canyon were primarily associated with zooplankton DVM, while spatial variations appeared to reflect the influence of the presence of the canyon and its morphology. The accumulation of organisms near the canyon slopes was evident in the U-shaped distribution of Sv as elevated values were found along both the NS and SS compared to the central axis of the canyon. This pattern suggests the interaction between zooplankton migration behavior and the steep topography of the BbC. Most zooplankton taxa undergo DVM within a 24 h cycle to reduce predation risk, typically descending to deeper waters during daylight and ascending to the surface at dusk (Forward, 1988; Hays, 2003; Bandara et al., 2021). In regions with abrupt or shallow topography, vertical migration may be blocked, resulting in the aggregation of organisms above topographic features, a mechanism called “topographic blocking”, often documented over seamounts (Aarflot et al., 2019; Mohn et al., 2021). Another form of topographic blocking occurs when horizontally transported zooplankton encounter steep bathymetric gradients, such as shallow banks (Isaacs and Schwartzlose, 1965) or the continental shelf break (Mackas et al., 1997). Submarine canyons have also been proposed as zones where such blocking can occur (Genin, 2004) due to their abrupt changes in depth. In our study, an accumulation of zooplankton was observed on the SS of the WT approximately 30 min after sunrise (Fig. S6, transect 8 at 08:30). This aggregation persisted throughout the day and was both shallow (∼ 75 m) and intense (> 60 dB re m−1) compared to the typical daytime Sv values within the canyon. These observations suggest that the daytime distribution of zooplankton in the WT is shaped by topographic blocking of their DVM.
Additionally, zooplankton DVM can interact with alternating vertical currents to promote or avoid retention. For example, some taxa exhibit vertical movements that interact with coastal wind-driven or tidal currents to either facilitate or avoid offshore advection (Castro et al., 1993; Miller and Shanks, 2004; Yannicelli et al., 2006b; Shanks et al., 2014; Meerhoff et al., 2015). In our observations, we frequently detected flow reversals with depth (see Fig. 4), as well as differences in Sv between vertical layers (see Figs. 9 and S6). The surface layer consistently exhibited an offshore flow, while currents in the mid-depth layer were predominantly positive, especially along the NS. Such opposing flow patterns could be exploited by zooplankton to promote retention.
The zooplankton community was composed of several taxa known for their strong migratory behavior (Mackas et al., 2005; dos Santos et al., 2008; Escribano et al., 2009; Bandara et al., 2021), such as decapod larvae, amphipods, and copepods. These organisms often utilize fluctuating vertical-flow regimes to avoid offshore advection by migrating between layers with contrasting current directions, a well-documented mechanism of zooplankton retention in tidal and estuarine systems (Castro et al., 1993; Hill, 1998; Poulin et al., 2002; Emsley et al., 2005; Kimmerer et al., 2014). This mechanism may have been present along the NS of the canyon. A migrating organism inhabiting the upper 30 m of the water column of the NS of the BbC would be subject to offshore transport. By migrating to deeper layers at sunrise, the organism could be transported onshore by the positive currents within the canyon. Upon ascending at dusk, it would again be exposed to offshore-flowing surface currents. Through this cycle, DVM behavior could facilitate retention within the canyon. This hypothesis might explain the elevated Sv values observed in the mid-depth layer of the NS during the daytime, whereas topographic blocking may account for the elevated Sv along the SS, particularly within the mid-depth layer of the WT.
4.2 Canyon-induced horizontal advection
Differences in zooplankton abundance between the NS and SS, observed in both acoustic and in situ sampling, were highly variable and alternating over time. These differences appeared to be associated with the contrasting cross-shore flows on either side of the canyon. Observational and numerical modeling studies have shown that submarine canyons can significantly influence and modify coastal circulation patterns (Sobarzo et al., 2016; Saldías and Allen, 2020; Figueroa et al., 2025). The presence of a shelf-incising canyon in a western continental margin often generates a flow dipole, characterized by an inshore flow along the downstream wall and an offshore flow along the upstream wall (Allen and Durrieu De Madron, 2009; Vergara et al., 2024). A recent study used high-resolution hydrodynamic simulations to investigate the influence of the BbC on the coastal circulation of the Arauco Gulf during upwelling and downwelling events (Vergara et al., 2024). The results revealed a persistent dipole in the mean cross-shore flow field. Specifically, the northern side of the canyon exhibited an onshore flow, while the southern area showed an offshore flow. Although this dipole pattern was present during both upwelling and downwelling conditions, the inshore flow was stronger under upwelling-favorable winds, resulting in a net onshore transport. The simulations also evidenced the advection of dense, deep water onto the shelf during upwelling, consistently with enhanced upwelling at the canyon head. Under downwelling conditions, a similar circulation dipole is formed, with minor differences in the magnitudes of offshore and onshore transports (Klinck, 1996; Spurgin and Allen, 2014; Figueroa et al., 2025). This dipole structure can trap particles within the canyon, promoting anticyclonic recirculation and particle retention for several days (Figueroa et al., 2025). Consequently, an upwelling precondition may favor the advection of offshore zooplankton into the canyon, whereas downwelling conditions may promote their retention through recirculation within the canyon.
The average cross-shore current revealed an inshore flow through the canyon, clearly tilted toward the NS in the ET, consistently with the theoretical dipole pattern. This flow configuration agrees with the upwelling-favorable wind conditions that preceded the study. For most of the sampling period, cross-shore flow in the 40–120 m depth layer was predominantly positive (onshore) along the NS of the ET and negative or alternating along the SS. The difference in the current magnitude and direction was more pronounced during the first 6 h of the study, coinciding with the shallowest pycnocline depths. The largest difference in Sv was also observed during this period, with Sv values being approximately ∼ 10 dB re m−1 higher along the NS compared to along the SS of the ET.
Thus, the inshore flow over the NS of the ET was associated with a shallower pycnocline, higher Sv, and increased zooplankton abundance. In the WT, located farther from the canyon head, the coherence between cross-shore flow and Sv patterns was less apparent. Nonetheless, a marked difference in Sv between the NS and SS of the WT emerged toward the end of the experiment, which was also reflected in the zooplankton samples, with higher abundances along the SS. During the final 6 h of the study, a strong offshore flow developed along the NS of the WT, potentially explaining the concurrent decline in Sv. Overall, the alternating Sv patterns and observed differences between the NS and SS across transects (see Figs. 6 and 9) throughout the 24 h cycle suggest that zooplankton may be unevenly transported within the canyon due to asymmetrical cross-shore circulation.
While our findings suggest that canyon-induced currents promote asymmetrical advection of zooplankton, they are based on a short (albeit intensive) observational period. Both flow and Sv variability were high, and the canyon's complex topography adds further challenges to interpretation. In the ET, alongshore flows between the NS and SS were highly correlated, likely due to the transect's proximity to the Biobio River mouth. In the WT, only the mid-depth layer showed correlated alongshore velocities between the NS and SS. For cross-shore velocities, only the deeper layer of the WT exhibited a correlation between the two sides of the canyon. Horizontal currents within the canyon not only differed between its walls but also shifted direction over timescales of less than a day. The BbC is a complex and elongated feature, with a smaller tributary canyon at its midpoint and pronounced changes in curvature from the continental slope to its head. This morphological complexity influences the spatial distribution of cross-shore transport and vertical velocities throughout the canyon (Vergara et al., 2024), potentially modifying the circulation patterns and particle retention mechanisms described in previous idealized numerical studies. These findings highlight the need for continued observational efforts to achieve a comprehensive understanding of the topographic upwelling driven by the canyon and to better resolve the net transport of zooplankton.
4.3 Ecological implications
Some submarine canyons are well-documented for their role in promoting zooplankton aggregations. Among the best-studied is Monterey Canyon in the California Current System, a known foraging site for large cetaceans (Schoenherr, 1991) and a critical krill habitat within an ecologically important canyon network (Santora et al., 2018). In contrast, submarine canyons within the HCS have only recently begun to receive detailed scientific attention. For example, earlier research on the Itata Canyon, located approximately 60 km north of the BbC, reported higher abundances of several crustacean larvae nearshore over the canyon, while offshore abundances dominated outside of it (Yannicelli et al., 2006a). Although the dynamics were not explicitly described, this pattern suggested potential inshore transport facilitated by the canyon. More recent work has proposed that a newly identified, relatively small canyon may explain the high concentration of whales and krill in northern Chile (Buchan et al., 2023). To our knowledge, this is the first study combining simultaneous observations of zooplankton aggregations and canyon-induced currents to investigate the physical mechanisms shaping zooplankton distributions.
Our findings suggest that the BbC plays a key role in shaping local zooplankton distributions through mechanisms such as asymmetric advection, topographic blocking, and particle retention processes influenced by the complex interplay of canyon morphology, hydrography, and circulation. These dynamics highlight the ecological importance of submarine canyons within the Humboldt Current upwelling system. However, due to the short temporal scale and the spatial limitations of our study, which focused primarily on intra-diurnal patterns and discrete sampling, further research is needed. Future efforts should incorporate long-term moorings to better capture the spatial and temporal variability of physical and biological processes. Expanding studies in underexplored, high-value regions is critical not only for advancing oceanographic knowledge but also for guiding conservation and management of these ecologically significant habitats.
We aimed to describe the spatio-temporal variability in zooplankton distribution and currents at the head of a long and narrow submarine canyon. Our results provide evidence that the canyon influenced zooplankton distribution and abundance within a day. The experiment was conducted following an event of upwelling-favorable winds under conditions of strong water column stratification driven by freshwater input from the Biobio River. Horizontal flows within the canyon were highly variable, with notable differences in flow velocity and direction between vertical layers and between the northern slope (NS) and the southern slope (SS). On average, an onshore flow was observed in both the western transect (WT) and the eastern transect (ET), with a tilt toward the NS in the ET. However, along-canyon currents frequently reversed direction at both slopes, resulting in low correlation between current velocities on either side of the canyon. Zooplankton abundance also varied spatially and temporally. At the beginning of the study, net abundances were higher along the NS than along the SS, but this pattern had reversed by the end of the experiment. A similar trend was observed in the acoustic backscatter (Sv), which was consistently higher near the canyon walls than at its center. In the WT, Sv was generally higher along the SS, while, in the ET, the NS showed higher Sv values. In the ET, peaks in Sv at the NS coincided in terms of time and depth with positive (onshore) flow, while the SS exhibited offshore flow, suggesting that asymmetric horizontal currents likely contributed to spatial differences in zooplankton distribution. We also found evidence of topographic blocking in the WT following sunrise. The interaction between zooplankton and opposing, alternating canyon flows may promote both their advection and retention. Overall, our findings demonstrate that submarine canyons are highly dynamic environments that significantly influence biological patterns at short timescales. These results highlight the importance of further studying submarine canyons, particularly in regions that provide essential ecosystem services.
The code used in this study built upon previous work by IPS and is constantly being updated. The code will be available upon reasonable request to the main author.
The datasets used in this study will be available upon reasonable request to the main author.
The supplement related to this article is available online at https://doi.org/10.5194/os-21-1833-2025-supplement.
MD-A: conceptualization, data curation, formal analysis, investigation, methodology, visualization, resources, writing (original draft preparation). MC: formal analysis, investigation, methodology, visualization, writing (review and editing). PF: formal analysis, data curation, visualization, writing (review and editing). LRC: resources, supervision, writing (review and editing). RR-B: investigation, supervision, writing (review and editing). IP-S: investigation, resources, supervision. OP: resources, writing (review and editing). GSS: conceptualization, investigation, methodology, funding acquisition, project administration, resources, supervision, writing (review and editing).
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
The authors are grateful to the R/V Kay-Kay II crew members, who supported the field sampling.
Macarena Díaz-Astudillo was supported by the ANID FONDECYT grant no. 3230183. Gonzalo S. Saldías was supported by FONDECYT grant no. 1220167. Iván Pérez-Santos was supported by FONDECYT grant nos. 1211037 and CIEP R20F002. Macarena Díaz-Astudillo, Gonzalo S. Saldías, Iván Pérez-Santos, and Leonardo R. Castro were partially supported by COPAS COASTAL grant no. ANID FB210021. This study was part of the field work funded by grant no. ANID AUB2200011 and FONDECYT grant no. 1220167.
This paper was edited by Karen J. Heywood and reviewed by Katherine L. Gallagher and one anonymous referee.
Aarflot, J. M., Aksnes, D. L., Opdal, A. F., Skjoldal, H. R., and Fiksen, Ø.: Caught in broad daylight: Topographic constraints of zooplankton depth distributions, Limnol. Oceanogr., 64, 849–859, https://doi.org/10.1002/lno.11079, 2019. a
Allen, S. E. and Durrieu de Madron, X.: A review of the role of submarine canyons in deep-ocean exchange with the shelf, Ocean Sci., 5, 607–620, https://doi.org/10.5194/os-5-607-2009, 2009. a, b, c, d
Amano, M., Aoki, K., Kobayashi, H., Minamikawa, S., Sato, K., and Kubodera, T.: Stereotypical diel movement and dive pattern of male sperm whales in a submarine canyon revealed by land-based and bio-logging surveys, Front. Mar. Sci., 10, 1150308, https://doi.org/10.3389/fmars.2023.1150308, 2023. a
Ancapichún, S. and Garcés-Vargas, J.: Variability of the Southeast Pacific Subtropical Anticyclone and its impact on sea surface temperature off north-central Chile, Cienc. Mar., 41, 1–20, https://doi.org/10.7773/cm.v41i1.2338, 2015. a
Aronés, K., Ayón, P., Hirche, H.-J., and Schwamborn, R.: Hydrographic structure and zooplankton abundance and diversity off Paita, northern Peru (1994 to 2004) — ENSO effects, trends and changes, J. Marine Syst., 78, 582–598, https://doi.org/10.1016/j.jmarsys.2009.01.002, 2009. a
Bandara, K., Varpe, Ø., Wijewardene, L., Tverberg, V., and Eiane, K.: Two hundred years of zooplankton vertical migration research, Biol. Rev., 96, 1547–1589, https://doi.org/10.1111/brv.12715, 2021. a, b
Barnes, S. L.: Applications of the Barnes objective analysis scheme. Part I: Effects of undersampling, wave position, and station randomness, J. Atmos. Ocean. Tech., 11, 1433–1448, 1994. a
Bozzano, R., Fanelli, E., Pensieri, S., Picco, P., and Schiano, M. E.: Temporal variations of zooplankton biomass in the Ligurian Sea inferred from long time series of ADCP data, Ocean Sci., 10, 93–105, https://doi.org/10.5194/os-10-93-2014, 2014. a
Brink, K. H.: The near-surface dynamics of coastal upwelling, Prog. Oceanogr., 12, 223–257, https://doi.org/10.1016/0079-6611(83)90009-5, 1983. a
Buchan, S. J., Ramos, M., Oyanadel, J., Santos-Carvallo, M., Bedriñana-Romano, L., Valladares, M., Maldonado, M., Astudillo, O., Sepúlveda, M., Pearce, S., and Olavarría, C.: Understanding the oceanographic dynamics of the Isla Chañaral baleen whale feeding ground, (Humboldt Archipelago, Northern Chile) to extend habitat protection, Front. Mar. Sci., 10, 1208262, https://doi.org/10.3389/fmars.2023.1208262, 2023. a, b
Cáceres, M., Valle-Levinson, A., Molinet, C., Castillo, M., Bello, M., and Moreno, C.: Lateral variability of flow over a sill in a channel of southern Chile, Ocean Dynam., 56, 352–359, https://doi.org/10.1007/s10236-006-0077-y, 2006. a
Castillo, M. I., Pizarro, O., Cifuentes, U., Ramirez, N., and Djurfeldt, L.: Subtidal dynamics in a deep fjord of southern Chile, Cont. Shelf Res., 49, 73–89, https://doi.org/10.1016/j.csr.2012.09.007, 2012. a
Castro, L. R., Bernal, P. A., and Troncoso, V. A.: Coastal intrusion of copepods: mechanisms and consequences on the population biology of Rhincalanus nasutus, J. Plankton Res., 15, 501–515, https://doi.org/10.1093/plankt/15.5.501, 1993. a, b
Cisewski, B., Hátún, H., Kristiansen, I., Hansen, B., Larsen, K. M. H., Eliasen, S. K., and Jacobsen, J. A.: Vertical Migration of Pelagic and Mesopelagic Scatterers From ADCP Backscatter Data in the Southern Norwegian Sea, Front. Mar. Sci., 7, 542386, https://doi.org/10.3389/fmars.2020.542386, 2021. a
Cisterna-Concha, A., Calderón-Quirgas, C., Silva-Andrades, F., Muñoz, R., and Norambuena, H. V.: Reencounter with the past: occurrence of sei whale (Balaenoptera borealis) in an old hunting area in the south-eastern Pacific Ocean, Nat. Conserv., 51, 1–12, https://doi.org/10.3897/natureconservation.51.95690, 2023. a
Clarke, J., Emmerson, L. M., and Otahal, P.: Envirnmental conditions and life history constraints determine foraging range in breeding Adélie penguins, Mar. Ecol.-Prog. Ser., 301, 247–261, 2006. a
Connolly, T. P. and Hickey, B. M.: Regional impact of submarine canyons during seasonal upwelling, J. Geophys. Res.-Oceans, 119, 953–975, https://doi.org/10.1002/2013JC009452.Received, 2014. a, b, c
Croll, D. A., Marinovic, B., Benson, S., Chavez, F. P., Black, N., Ternullo, R., and Tershy, B. R.: From wind to whales: Trophic links in a coastal upwelling system, Mar. Ecol.-Prog. Ser., 289, 117–130, https://doi.org/10.3354/meps289117, 2005. a
De Leo, F. C., Drazen, J. C., Vetter, E. W., Rowden, A. A., and Smith, C. R.: The effects of submarine canyons and the oxygen minimum zone on deep-sea fish assemblages off Hawai'i, Deep-Sea Res. Pt. I, 64, 54–70, https://doi.org/10.1016/j.dsr.2012.01.014, 2012. a
Deines, K. L.: Backscatter estimation using Broadband acoustic Doppler current profilers, Proceedings of the IEEE Sixth Working Conference on Current Measurement (Cat. No.99CH36331), 249–253, https://doi.org/10.1109/CCM.1999.755249, 1999. a
Díaz-Astudillo, M., Riquelme-Bugueño, R., Bernard, K. S., Saldías, G. S., Rivera, R., and Letelier, J.: Disentangling species-specific krill responses to local oceanography and predator's biomass: The case of the Humboldt krill and the Peruvian anchovy, Front. Mar. Sci., 9, 1–19, https://doi.org/10.3389/fmars.2022.979984, 2022. a
Díaz-Astudillo, M., Riquelme-Bugueño, R., Saldías, G. S., and Letelier, J.: Mesoscale and climate environmental variability drive krill community changes in the Humboldt Current System, J. Marine Syst., 245, 103998, https://doi.org/10.1016/j.jmarsys.2024.103998, 2024. a
dos Santos, A., Santos, A., Conway, D., Bartilotti, C., Lourenço, P., and Queiroga, H.: Diel vertical migration of decapod larvae in the Portuguese coastal upwelling ecosystem: implications for offshore transport, Mar. Ecol.-Prog. Ser., 359, 171–183, https://doi.org/10.3354/meps07341, 2008. a
Dwinovantyo, A., Manik, H. M., Prartono, T., Susilohadi, S., and Mukai, T.: Variation of Zooplankton mean volume backscattering strength from Moored and Mobile ADCP instruments for diel vertical migration observation, Appl. Sci., 9, 1851, https://doi.org/10.3390/app9091851, 2019. a
Emsley, S. M., Tarling, G. A., and Burrows, M. T.: The effect of vertical migration strategy on retention and dispersion in the Irish Sea during spring–summer, Fish. Oceanogr., 14, 161–174, https://doi.org/10.1111/j.1365-2419.2005.00327.x, 2005. a
Escribano, R., Hidalgo, P., and Krautz, C.: Zooplankton associated with the oxygen minimum zone system in the northern upwelling region of Chile during March 2000, Deep-Sea Res. Pt. II, 56, 1083–1094, https://doi.org/10.1016/j.dsr2.2008.09.009, 2009. a
Escribano, R., Hidalgo, P., Fuentes, M., and Donoso, K.: Zooplankton time series in the coastal zone off Chile: Variation in upwelling and responses of the copepod community, Prog. Oceanogr., 97-100, 174–186, https://doi.org/10.1016/j.pocean.2011.11.006, 2012. a
Fernandez-Arcaya, U., Ramirez-Llodra, E., Aguzzi, J., Allcock, A. L., Davies, J. S., Dissanayake, A., Harris, P., Howell, K., Huvenne, V. A., Macmillan-Lawler, M., Martín, J., Menot, L., Nizinski, M., Puig, P., Rowden, A. A., Sanchez, F., and Van den Beld, I. M.: Ecological role of submarine canyonsand need for canyon conservation: A review, Front. Mar. Sci., 4, 1–26, https://doi.org/10.3389/fmars.2017.00005, 2017. a
Fielding, S., Griffiths, G., and Roe, H.: The biological validation of ADCP acoustic backscatter through direct comparison with net samples and model predictions based on acoustic-scattering models, ICES J. Mar. Sci., 61, 184–200, https://doi.org/10.1016/j.icesjms.2003.10.011, 2004. a
Figueroa, P. A., Saldías, G. S., and Allen, S. E.: The influence of a submarine canyon on the wind-driven downwelling circulation over the continental shelf, Ocean Sci., 21, 643–659, https://doi.org/10.5194/os-21-643-2025, 2025. a, b, c
Forward, R. B. J.: Diel vertical migration: zooplankton photobiology and behaviour, Oceanogr. Mar. Biol., 26, 361–393, 1988. a
Genin, A.: Bio-physical coupling in the formation of zooplankton and fish aggregations over abrupt topographies, J. Marine Syst., 50, 3–20, https://doi.org/10.1016/j.jmarsys.2003.10.008, 2004. a, b, c, d, e
Harris, P. T. and Whiteway, T.: Global distribution of large submarine canyons: Geomorphic differences between active and passive continental margins, Mar. Geol., 285, 69–86, https://doi.org/10.1016/j.margeo.2011.05.008, 2011. a
Hays, G. C.: A review of the adaptive significance and ecosystem consequences of zooplankton diel vertical migrations, Hydrobiologia, 503, 163–170, https://doi.org/10.1023/B:HYDR.0000008476.23617.b0, 2003. a
Hersbach, H., Bell, B., Berrisford, P., Hirahara, S., Horányi, A., Muñoz‐Sabater, J., Nicolas, J., Peubey, C., Radu, R., Schepers, D., Simmons, A., Soci, C., Abdalla, S., Abellan, X., Balsamo, G., Bechtold, P., Biavati, G., Bidlot, J., Bonavita, M., De Chiara, G., Dahlgren, P., Dee, D., Diamantakis, M., Dragani, R., Flemming, J., Forbes, R., Fuentes, M., Geer, A., Haimberger, L., Healy, S., Hogan, R. J., Hólm, E., Janisková, M., Keeley, S., Laloyaux, P., Lopez, P., Lupu, C., Radnoti, G., de Rosnay, P., Rozum, I., Vamborg, F., Villaume, S., and Thépaut, J.-N.: The ERA5 global reanalysis, Q. J. Roy. Meteor. Soc., 146, 1999–2049, https://doi.org/10.1002/qj.3803, 2020. a
Hill, a. E.: Diel vertical migration in stratified tidal flows: Implications for plankton dispersal, J. Marine Res., 56, 1069–1096, https://doi.org/10.1357/002224098765173464, 1998. a
Hudson, K., Oliver, M. J., Kohut, J., Cohen, J. H., Dinniman, M. S., Klinck, J. M., Reiss, C. S., Cutter, G. R., Statscewich, H., Bernard, K. S., and Fraser, W.: Subsurface Eddy Facilitates Retention of Simulated Diel Vertical Migrators in a Biological Hotspot, J. Geophys. Res.-Oceans, 127, 1–15, https://doi.org/10.1029/2021JC017482, 2022a. a
Hudson, K., Oliver, M. J., Kohut, J., Dinniman, M. S., Klinck, J. M., Cimino, M. A., Bernard, K. S., Statscewich, H., and Fraser, W.: A subsurface eddy associated with a submarine canyon increases availability and delivery of simulated Antarctic krill to penguin foraging regions, Mar. Ecol.-Prog. Ser., 702, 105–122, https://doi.org/10.3354/meps14211, 2022b. a, b, c
Isaacs, J. D. and Schwartzlose, R. A.: Migrant Sound Scatterers: Interaction with the Sea Floor, Science, 150, 1810–1813, https://doi.org/10.1126/science.150.3705.1810, 1965. a
Jiang, S., Dickey, T. D., Steinberg, D. K., and Madin, L. P.: Temporal variability of zooplankton biomass from ADCP backscatter time series data at the Bermuda Testbed Mooring site, Deep-Sea Res. Pt. I, 54, 608–636, https://doi.org/10.1016/j.dsr.2006.12.011, 2007. a
Joyce, T. M.: On In Situ “Calibration” of Shipboard ADCPs, J. Atmos. Ocean. Tech., 6, 169–172, https://doi.org/10.1175/1520-0426(1989)006<0169:OISOSA>2.0.CO;2, 1989. a
Kimmerer, W. J., Gross, E. S., and MacWilliams, M. L.: Tidal migration and retention of estuarine zooplankton investigated using a particle‐tracking model, Limnol. Oceanogr., 59, 901–916, https://doi.org/10.4319/lo.2014.59.3.0901, 2014. a
Klinck, J. M.: Circulation near submarine canyons: A modeling study, J. Geophys. Res.-Oceans, 101, 1211–1223, 1996. a
Landaeta, M. F. and Castro, L. R.: Spring spawning and early nursery zone of the mesopelagic fish Maurolicus parvipinnis at the coastal upwelling zone off Talcahuano, central Chile , Mar. Ecol.-Prog. Ser., 226, 179–191, 2002. a
Landaeta, M. F., Veas, R., Letelier, J., and Castro, L. R.: Larval fish assemblages off central Chile upwelling ecosystem, Rev. Biol. Mar. Oceanog., 43, 569–584, https://doi.org/10.4067/s0718-19572008000300016, 2008. a
Lee, K., Mukai, T., Kang, D., and Iida, K.: Application of acoustic Doppler current profiler combined with a scientific echo sounder for krill Euphausia pacifica density estimation, Fish. Sci., 70, 1051–1060, https://doi.org/10.1111/j.1444-2906.2004.00905.x, 2004. a
Lwiza, K., Bowers, D., and Simpson, J.: Residual and tidal flow at a tidal mixing front in the North Sea, Cont. Shelf Res., 11, 1379–1395, https://doi.org/10.1016/0278-4343(91)90041-4, 1991. a, b
Mackas, D., Tsurumi, M., Galbraith, M., and Yelland, D.: Zooplankton distribution and dynamics in a North Pacific Eddy of coastal origin: II. Mechanisms of eddy colonization by and retention of offshore species, Deep-Sea Res. Pt. II, 52, 1011–1035, https://doi.org/10.1016/j.dsr2.2005.02.008, 2005. a
Mackas, D. L., Kieser, R., Saunders, M., Yelland, D. R., Brown, R. M., and Moore, D. F.: Aggregation of euphausiids and Pacific hake (Merluccius productus) along the outer continental shelf off Vancouver Island, Can. J. Fish. Aquat. Sci., 54, 2080–2096, https://doi.org/10.1139/f97-113, 1997. a
Masotti, I., Aparicio-Rizzo, P., Yevenes, M. A., Garreaud, R., Belmar, L., and Farías, L.: The Influence of River Discharge on Nutrient Export and Phytoplankton Biomass Off the Central Chile Coast (33°–37° S): Seasonal Cycle and Interannual Variability, Front. Mar. Sci., 5, 423, https://doi.org/10.3389/fmars.2018.00423, 2018. a
Medellín-Mora, J., Escribano, R., and Schneider, W.: Community response of zooplankton to oceanographic changes (2002–2012) in the central/southern upwelling system of Chile, Prog. Oceanogr., 142, 17–29, https://doi.org/10.1016/j.pocean.2016.01.005, 2016. a
Meerhoff, E., Tapia, F. J., Sobarzo, M., and Castro, L.: Influence of estuarine and secondary circulation on crustacean larval fluxes: a case study from a Patagonian fjord, J. Plankton Res., 37, 168–182, https://doi.org/10.1093/plankt/fbu106, 2015. a
Miller, J. A. and Shanks, A. L.: Ocean-estuary coupling in the Oregon upwelling region: abundance and transport of juvenile fish and of crab megalopae, Mar. Ecol.-Prog. Ser., 271, 267–279, 2004. a
Mohn, C., White, M., Denda, A., Erofeeva, S., Springer, B., Turnewitsch, R., and Christiansen, B.: Dynamics of currents and biological scattering layers around Senghor Seamount, a shallow seamount inside a tropical Northeast Atlantic eddy corridor, Deep-Sea Res. Pt. I, 171, 103497, https://doi.org/10.1016/j.dsr.2021.103497, 2021. a
Moors-Murphy, H. B.: Submarine canyons as important habitat for cetaceans, with special reference to the Gully: A review, Deep-Sea Res. Pt. II, 104, 6–19, https://doi.org/10.1016/j.dsr2.2013.12.016, 2014. a
Morales, C. E., González, H. E., Hormazabal, S. E., Yuras, G., Letelier, J., and Castro, L. R.: The distribution of chlorophyll-a and dominant planktonic components in the coastal transition zone off Concepción, central Chile, during different oceanographic conditions, Prog. Oceanogr., 75, 452–469, https://doi.org/10.1016/j.pocean.2007.08.026, 2007. a
Morales, C. E., Loreto Torreblanca, M., Hormazabal, S., Correa-Ramírez, M., Nuñez, S., and Hidalgo, P.: Mesoscale structure of copepod assemblages in the coastal transition zone and oceanic waters off central-southern Chile, Prog. Oceanogr., 84, 158–173, https://doi.org/10.1016/j.pocean.2009.12.001, 2010. a
Mullison, J.: Backscatter Estimation Using Broadband Acoustic Dopplet Current Profilers – Updated, Tech. rep., Teledyne RD Instruments, 2017. a
Pavez, M. A., Landaeta, M. F., Castro, L. R., and Schneider, W.: Distribution of carnivorous gelatinous zooplankton in the upwelling zone off central Chile (austral spring 2001), J. Plankton Res., 32, 1051–1065, https://doi.org/10.1093/plankt/fbq029, 2010. a
Pollard, R. and Read, J.: A Method for Calibrating Shipmounted Acoustic Doppler Profilers and the Limitations of Gyro Compasses, J. Atmos. Ocean. Tech., 6, 859–865, https://doi.org/10.1175/1520-0426(1989)006<0859:AMFCSA>2.0.CO;2, 1989. a
Poulin, E., Palma, A. T., Leiva, G., Narvaez, D., Pacheco, R., Navarrete, S. A., and Castilla, J. C.: Avoiding offshore transport of competent larvae during upwelling events: The case of the gastropod Concholepas concholepas in Central Chile, Limnol. Oceanogr., 47, 1248–1255, https://doi.org/10.4319/lo.2002.47.4.1248, 2002. a
Prairie, J. C., Sutherland, K. R., Nickols, K. J., and Kaltenberg, A. M.: Biophysical interactions in the plankton: A cross-scale review, Limnol. Oceanogr., 2, 121–145, https://doi.org/10.1215/21573689-1964713, 2012. a
Riquelme-Bugueño, R., Núñez, S., Jorquera, E., Valenzuela, L., Escribano, R., and Hormazabal, S. E.: The influence of upwelling variation on the spatially-structured euphausiid community off central-southern Chile in 2007-2008, Prog. Oceanogr., 92–95, 146–165, https://doi.org/10.1016/j.pocean.2011.07.003, 2012. a
Riquelme‐Bugueño, R., Correa‐Ramírez, M., Escribano, R., Núñez, S., and Hormazábal, S.: Mesoscale variability in the habitat of the Humboldt Current krill, spring 2007, J. Geophys. Res.-Oceans, 120, 2769–2783, https://doi.org/10.1002/2014JC010460, 2015. a
Rodrigo, C.: Cañones submarinos en el margen continental chileno, in: Geología Marina de Chile, edited by: Díaz, J. and Frutos, J., 32–35, Comité Oceanográfico Nacional CONA, Valparaíso, Chile, ISBN 978-956-235-026-6, 2010. a
Rojas, C. M., Saldías, G. S., Flores, R. P., Vásquez, S. I., Salas, C., and Vargas, C. A.: A modeling study of hydrographic and flow variability along the river-influenced coastal ocean off central Chile, Ocean Model., 181, 102155, https://doi.org/10.1016/j.ocemod.2022.102155, 2023. a
Saldías, G. S. and Allen, S. E.: The influence of a submarine canyon on the circulation and cross-shore exchanges around an upwelling front, J. Phys. Oceanogr., 50, 1677–1698, https://doi.org/10.1175/JPO-D-19-0130.1, 2020. a, b
Saldías, G. S., Sobarzo, M., Largier, J., Moffat, C., and Letelier, R.: Seasonal variability of turbid river plumes off central Chile based on high-resolution MODIS imagery, Remote Sens. Environ., 123, 220–233, https://doi.org/10.1016/j.rse.2012.03.010, 2012. a
Saldías, G. S., Largier, J. L., Mendes, R., Pérez-Santos, I., Vargas, C. A., and Sobarzo, M.: Satellite-measured interannual variability of turbid river plumes off central-southern Chile: Spatial patterns and the influence of climate variability, Prog. Oceanogr., 146, 212–222, https://doi.org/10.1016/j.pocean.2016.07.007, 2016. a
Saldías, G. S., Ramos-Musalem, K., and Allen, S. E.: Circulation and Upwelling Induced by Coastal Trapped Waves Over a Submarine Canyon in an Idealized Eastern Boundary Margin, Geophys. Res. Lett., 48, 1–10, https://doi.org/10.1029/2021GL093548, 2021. a
Salgado Kent, C., Bouchet, P., Wellard, R., Parnum, I., Fouda, L., and Erbe, C.: Seasonal productivity drives aggregations of killer whales and other cetaceans over submarine canyons of the Bremer Sub-Basin, south-western Australia, Aust. Mammal., 43, 168–178, https://doi.org/10.1071/AM19058, 2021. a
Santora, J. A. and Reiss, C. S.: Geospatial variability of krill and top predators within an Antarctic submarine canyon system, Mar. Biol., 158, 2527–2540, https://doi.org/10.1007/s00227-011-1753-0, 2011. a
Santora, J. A., Zeno, R., Dorman, J. G., and Sydeman, W. J.: Submarine canyons represent an essential habitat network for krill hotspots in a Large Marine Ecosystem, Sci. Rep., 8, 1–9, https://doi.org/10.1038/s41598-018-25742-9, 2018. a, b
Saunders, B. J., Galaiduk, R., Inostroza, K., Myers, E. M. V., Goetze, J. S., Westera, M., Twomey, L., McCorry, D., and Harvey, E. S.: Quantifying Patterns in Fish Assemblages and Habitat Use Along a Deep Submarine Canyon-Valley Feature Using a Remotely Operated Vehicle, Front. Mar. Sci., 8, 608665, https://doi.org/10.3389/fmars.2021.608665, 2021. a
Schoenherr, J. R.: Blue whales feeding on high concentrations of euphausiids around Monterey Submarine Canyon, Can. J. Zool., 69, 583–594, https://doi.org/10.1139/z91-088, 1991. a, b
Schofield, O., Ducklow, H., Bernard, K., Doney, S., Patterson-Fraser, D., Gorman, K., Martinson, D., Meredith, M., Saba, G., John, S. S., Steinberg, D. K., and Fraser, W.: Penguin biogeography along the West Antarctic Peninsula: Testing the canyon hypothesis with palmer LTER observations, Oceanography, 26, 204–206, https://doi.org/10.5670/oceanog.2013.63, 2013. a
Shanks, A., Morgan, S., Macmahan, J., Reniers, A., Jarvis, M., Brown, J., Fujimura, A., and Griesemer, C.: Onshore transport of plankton by internal tides and upwelling-relaxation events, Mar. Ecol.-Prog. Ser., 502, 39–51, 2014. a
Silva, M. V. B. and Araújo, T. C. M.: The South American Advances in Submarine Canyons Studies and Their Link to the Ocean Decade, Front. Mar. Sci., 8, 764029, https://doi.org/10.3389/fmars.2021.764029, 2021. a
Sobarzo, M., Bravo, L., Donoso, D., Garcés-Vargas, J., and Schneider, W.: Coastal upwelling and seasonal cycles that influence the water column over the continental shelf off central Chile, Prog. Oceanogr., 75, 363–382, https://doi.org/10.1016/j.pocean.2007.08.022, 2007. a, b, c
Sobarzo, M., Saldías, G. S., Tapia, F. J., Bravo, L., and Moffat, C.: On subsurface cooling associated with the Biobio River Canyon (Chile), J. Geophys. Res., 121, 4568–4584, https://doi.org/10.1002/2016JC011796, 2016. a, b, c, d
Soto, L., Saldías, G. S., Pavez, R., González, M., Camaño, C., Valenzuela, C., Núñez, S., González, C., San Martín, R., Zúñiga, A., Vásquez, S., and Arteaga, M.: Informe Final Proyecto FIPA No. 2019-22, Abundancia y distribución de moluscos, crustáceos y peces costeros de importancia comercial y su relación con la variabilidad de precipitaciones y caudal de ríos en Chile, Tech. rep., Fondo de Investigación Pesquera, 2022. a
Spurgin, J. M. and Allen, S. E.: Flow dynamics around downwelling submarine canyons, Ocean Sci., 10, 799–819, https://doi.org/10.5194/os-10-799-2014, 2014. a
Trump, C. L. and Marmorino, G. O.: Calibrating a Gyrocompass Using ADCP and DGPS Data, J. Atmos. Ocean. Tech., 14, 211–214, https://doi.org/10.1175/1520-0426(1997)014<0211:CAGUAA>2.0.CO;2, 1997. a
Vergara, O. A., Saldías, G., Salas, C., and Figueroa, P.: The influence of the Biobio submarine canyon on the circulation and coastal upwelling/downwelling off central Chile, Cont. Shelf Res., 282, 105335, https://doi.org/10.1016/j.csr.2024.105335, 2024. a, b, c, d, e, f
Vindeirinho, C.: Water properties, currents and zooplankton distribution over a submarine canyon under upwelling-favorable conditions, Master's thesis, The University of British Columbia, https://doi.org/10.14288/1.0053340, 1998. a
Wang, H., Gong, D., Friedrichs, M. A., Harris, C. K., Miles, T., Yu, H. C., and Zhang, Y.: A Cycle of Wind-Driven Canyon Upwelling and Downwelling at Wilmington Canyon and the Evolution of Canyon-Upwelled Dense Water on the MAB Shelf, Front. Mar. Sci., 9, 1–16, https://doi.org/10.3389/fmars.2022.866075, 2022. a
Yannicelli, B., Castro, L., Schneider, W., and Sobarzo, M.: Crustacean larvae distribution in the coastal upwelling zone off Central Chile, Mar. Ecol.-Prog. Ser., 319, 175–189, https://doi.org/10.3354/meps319175, 2006a. a, b
Yannicelli, B., Castro, L. R., Valle-Levinson, A., Atkinson, L., and Figueroa, D.: Vertical distribution of decapod larvae in the entrance of an equatorward facing bay of central Chile: implications for transport, J. Plankton Res., 28, 19–37, https://doi.org/10.1093/plankt/fbi098, 2006b. a, b