the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 24 Jul 2025
| 24 Jul 2025
Sargassum spp. accumulation and transport by mesoscale eddies
Rosmery Sosa-Gutierrez
Leo Berline
The proliferation of pelagic Sargassum spp. (Sargassum) in the tropical Atlantic has significant ecological and socioeconomic impacts. While large-scale ocean circulation patterns influence the basin-scale distribution of Sargassum, the role of mesoscale eddies in their local accumulation and transport has not been quantitatively assessed so far. This study investigates the relationship between mesoscale eddies and Sargassum dynamics using satellite observations. By analyzing 13 years of remote sensing observations, we demonstrate that both cyclonic and anticyclonic long-lived mesoscale eddies can trap and transport Sargassum. However, results show that, in cyclonic eddies, Sargassum cover is higher and tends to accumulate during its lifetime, while within anticyclonic eddies the Sargassum cover is usually weaker and tends to decrease. These findings align with recent studies highlighting the role of eddies in shaping the distribution of floating debris and provide an important observational basis for the development of Sargassum drift models.
- Article
(1540 KB) - Full-text XML
- BibTeX
- EndNote




Prior to 2010, the two holopelagic Sargassum species, Sargassum natans and Sargassum fluitans (Sargassum hereafter), were found primarily in the Sargasso Sea and the northwestern tropical Atlantic (Gower and King, 2011). However, since 2010, these two pelagic Sargassum species have expanded their presence to the tropical Atlantic (<20° N), from the coasts of the Lesser Antilles, Central America, and Brazil to western Africa (Gower et al., 2013; Wang et al., 2019). This expansion has led to stranding events and large accumulations, resulting in significant economic and environmental damage in the coastal areas of the tropical Atlantic (e.g., Rodríguez-Martínez et al., 2024; Van Tussenbroek et al., 2017; Hendy et al., 2021; Antonio-Martínez et al., 2020; Rosellón-Druker et al., 2023).
Sargassum remains afloat in the upper ocean due to its gas-filled bladders, making it highly responsive to both surface currents and wind. The dynamics of the upper ocean play a critical role in the formation of Sargassum accumulations, which can occur across a wide range of spatial scales (see Ody et al., 2019). At smaller scales, on the order of tens of meters, accumulations are typically driven by Langmuir circulation (Langmuir, 1938). At larger scales, reaching hundreds of kilometers, mesoscale and submesoscale frontal dynamics become dominant (Gower et al., 2013; Zhong et al., 2012). In particular, convergence zones associated with submesoscale dynamics have been shown to concentrate buoyant material and to structure ecosystems (D'Asaro et al., 2018; Esposito et al., 2021; Lévy et al., 2018). However, the role of mesoscale eddies in the accumulation and transport of Sargassum remains uncertain. Early theoretical and experimental work by Provenzale (1999) suggested that heavy impurities can be concentrated in the cores of anticyclonic vortices. Beron-Vera et al. (2015) provided both theoretical justification and numerical evidence for a more general principle governing the behavior of inertial particles near quasi-geostrophic eddies: anticyclonic (cyclonic) eddies tend to attract heavy (light) particles and repel light (heavy) ones. More recent advances, incorporating wind drag and elastic forces into the Maxey–Riley equations, have shown that these additional factors can have opposing influences sometimes favoring accumulation in anticyclones rather than cyclones (Beron-Vera, 2021). Observational studies also present a mixed picture. For instance, limited in situ measurements have shown micro-plastic accumulation within anticyclonic eddies (Brach et al., 2018), while a more systematic study by Vic et al. (2022) demonstrated a tendency for drogued drifters to accumulate in cyclonic structures in the North Atlantic. These contrasting findings highlight the complexity and context dependence of floating object dynamics in mesoscale eddies. The accumulation and transport behavior likely depend on the specific properties of the objects in question – especially their buoyancy and windage.
With respect to Sargassum, a few case studies have shown that mesoscale eddies can transport Sargassum. For example, Andrade-Canto et al. (2022), using 8 years of satellite altimetry in the eastern Caribbean Sea, showed that mesoscale eddies (both cyclonic and anticyclonic) can transport Sargassum, and more recently Sun et al. (2024) presented some illustrative cases of accumulation in eddies. However, there has been no systematic assessment of the transport and organization of Sargassum by these eddies. We fill this gap through a systematic analysis of Sargassum distribution by combining eddy-tracking techniques from altimetry (Chaigneau et al., 2009; Pegliasco et al., 2015; Sosa-Gutiérrez et al., 2020) and long-term Sargassum detection from the MODIS ocean color sensor (Descloitres et al., 2021) for the last 13 years. There are several known eddy detection and tracking algorithms based on different methods, such as Okubo–Weiss (Okubo, 1970; Weiss, 1991), vector geometry (Nencioli et al., 2010), winding angle (Chaigneau et al., 2009; Chen et al., 2011), and, recently, geodesic detection (Andrade-Canto et al., 2022). In this work, we opted for an Eulerian detection method due to its relative ease of implementation. This paper is organized as follows. Section 2 describes the data and methods used. The main characteristics of Sargassum cover in cyclonic and anticyclonic eddies are presented in the Results section (Sect. 3). Finally, in Sect. 4, we present the summary and discussion of our results.
Sargassum detections were obtained from the SAREDA database (Sargassum Evolving Distributions in the Atlantic, Descloitres et al., 2021). This product estimates Sargassum cover by computing the Alternative Floating Algae Index (AFAI; Wang and Hu, 2016) from ocean color acquisitions using MODIS, which operates aboard the Aqua and Terra satellites. The AFAI, computed using the processing described in Descloitres et al. (2021), was converted to fractional cover (FC), which represents the proportion of Sargassum cover in each pixel. Daily FC at 1 km from the SAREDA database was aggregated on a regular grid of 0.25° (∼25 km) horizontal resolution. These daily MODIS images have significant cloud coverage compared to multi-sensor products (e.g., Sun et al., 2024) but have the advantage of being a homogeneous series over the last 13 years. They have already allowed the tracking of a rapid decrease in Sargassum coverage in the lee of tropical cyclones (Sosa-Gutierrez et al., 2022). They therefore seem well suited to the mesoscale compositing proposed in the present study. Sargassum biomass is obtained from Sargassum cover by considering an average density of 3.34 kg m−2 for pure Sargassum patches as estimated by Wang et al. (2018).
Mesoscale cyclonic and anticyclonic eddies in the tropical Atlantic were identified from 2011 to 2023 using daily absolute dynamic topography (ADT) maps at a horizontal resolution of 25 km (EU Copernicus Marine Service Product, 2024), which are distributed by the Copernicus Marine and Environment Monitoring Service (CMEMS; https://www.copernicus.eu/en, last access: November 2024). The mesoscale eddies were detected using a method described by Chaigneau et al. (2009), with a slight modification of the methodology outlined by Sosa-Gutiérrez et al. (2020). While the original method identifies the outermost closed contour of sea level anomalies (SLAs) for each anticyclonic and cyclonic eddy center (SLA maxima and minima, respectively) as eddy edges, the modification involves considering the outermost closed SLA where the averaged azimuthal velocity is maximum at the periphery of the eddy. The eddy-tracking algorithm, described in Pegliasco et al. (2015), involves calculating the paths of eddies by intersecting them with daily maps. For this study, only mesoscale eddies with radii larger than 40 km and lifetimes longer than 30 d were retained.
An illustration of 7 d averaged Sargassum cover centered on 28 June 2021, together with superimposed eddy detections for the same day, is given in Fig. 1. This figure illustrates that the Sargassum distribution is partially structured by mesoscale eddies. To provide a systematic and quantitative view of this organization by mesoscale eddies, we composited the Sargassum coverage by selecting eddies under the following conditions: (i) the detected eddy lasts at least 30 d, (ii) during its lifetime the eddy has more than 25 % of unclouded ocean color observations, and (iii) the Sargassum presence is detected at least once during the eddy's lifetime. This results in 471 trajectories of anticyclonic eddies (AEs) and 628 trajectories of cyclonic eddies (CEs) between 2011 and 2023 (Fig. 2a and b, respectively). The trajectory analysis shows that AEs are predominantly found in the path of the western boundary current from the North Brazil Current (NBC) to the Loop Current, in agreement with the known distribution of large anticyclonic eddies in this region (e.g., Richardson, 2005; Jouanno et al., 2008). In contrast, CEs are more homogenously distributed in the central Atlantic and the Caribbean Sea. The mean lifetimes of AEs that captured Sargassum were 69±40 d (mean ± standard deviation) and 65±37 for CE; the longest-lived AEs lasted 277 and 269 d for a CE (Fig. 2c and d).
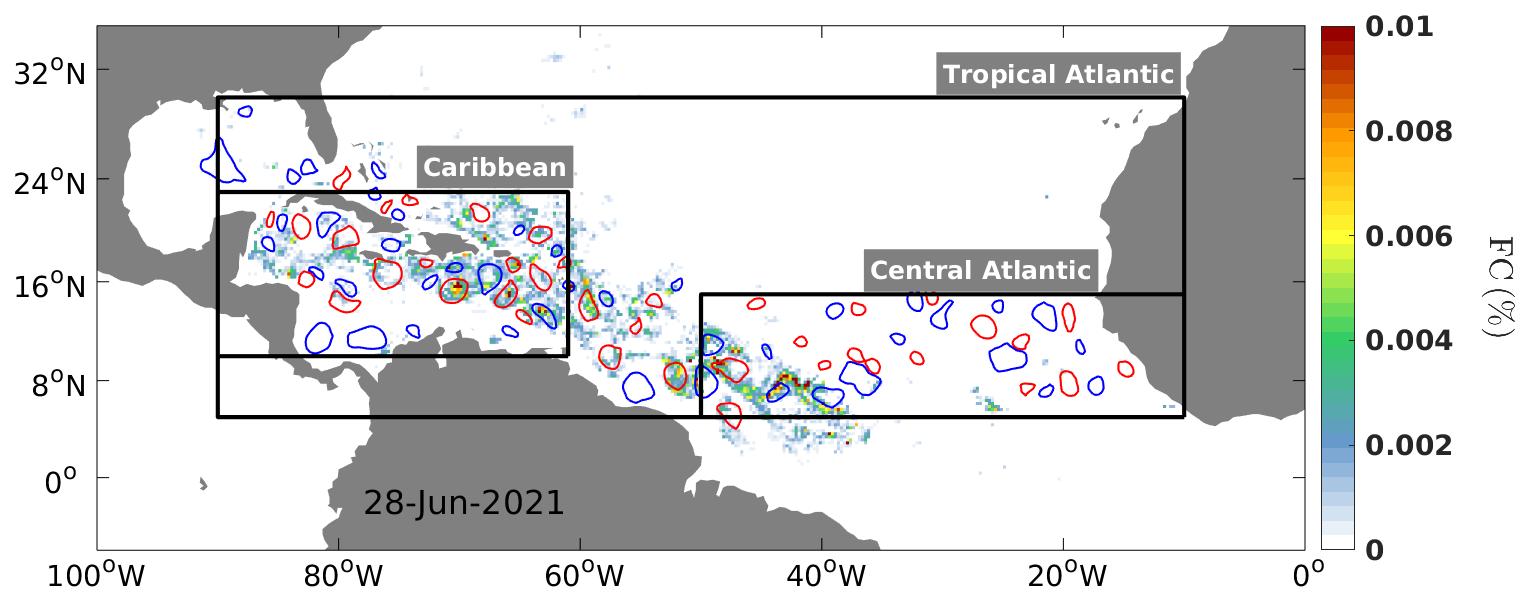
Figure 1Distribution of Sargassum cover (%) obtained from MODIS and mesoscale eddy contours detected with the CMEMS ADT product for 28 June 2021. Sargassum biomass was averaged over a 7 d period centered on 28 June and coarsened to a 0.25° regular grid. Red contours represent anticyclonic eddies, and blue contours represent cyclonic eddies. Only eddies with non-zero FCs during their lifetimes are shown.
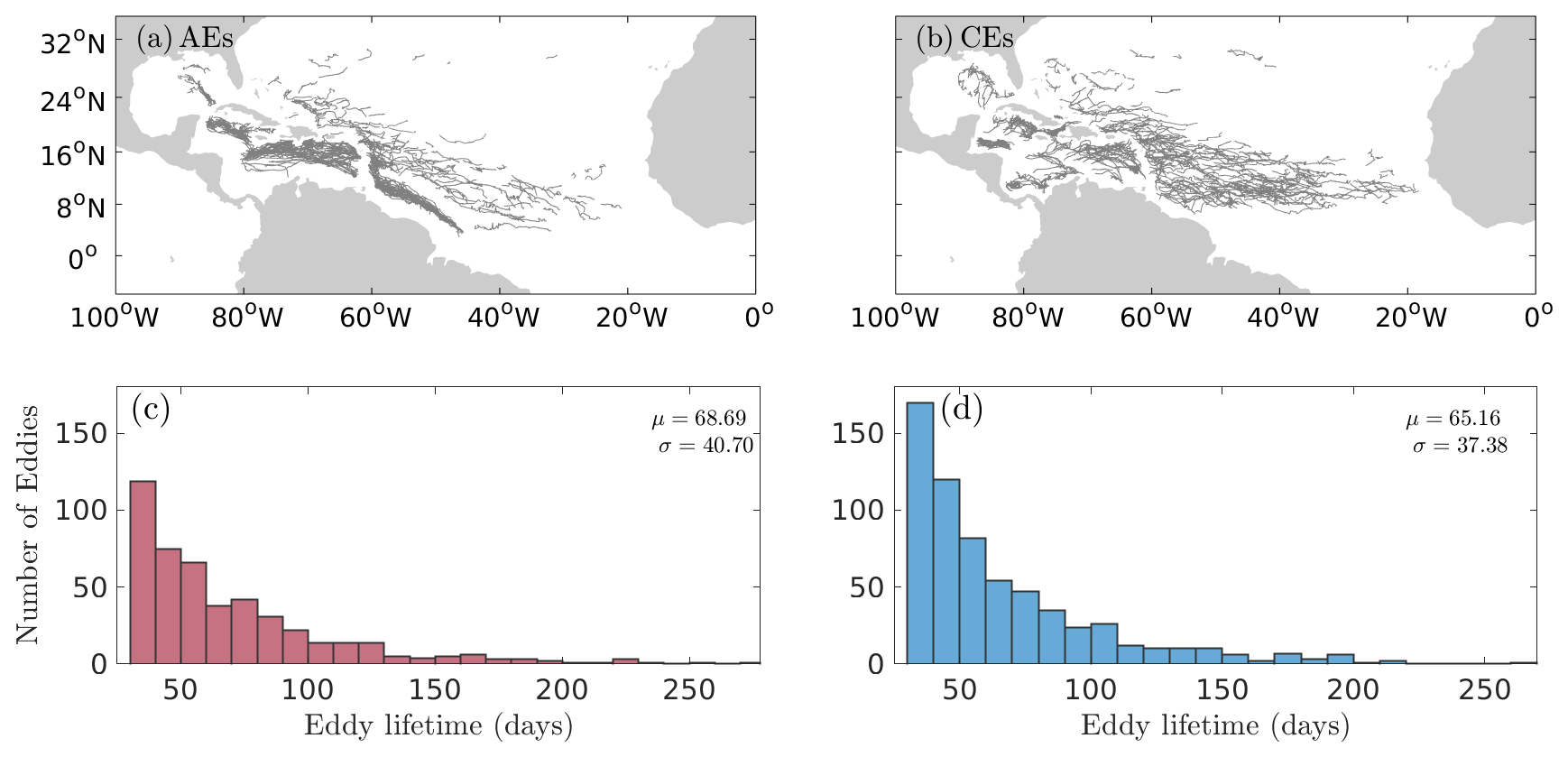
Figure 2Eddy tracks for the period 2011–2023 with (a) trajectories of AEs and (b) trajectories of CEs that last at least 30 d and have seen Sargassum during their lifetimes. Panels (c) and (d) show histograms of the total number of AEs (red) and CEs (blue) with respect to eddy lifetimes (d).
The Sargassum cover within CEs and AEs and their neighborhood was averaged for each day within the period 2011–2023 for the entire tropical Atlantic (5–30° N, 10–90° W) and in two regions where mesoscale activity and Sargassum presence are significant: the Caribbean Sea (10–23° N, 61–90° W) and the central Atlantic (5–15° N, 10–50° W). To construct the composite distribution, each eddy was normalized by its radius, where a radius of ±1 corresponds to the eddy periphery, 0 corresponds to the eddy center, and a radius larger than +1 or lower than −1 corresponds to the region surrounding the eddy (Fig. 3). This normalization allowed us to account for eddies of different sizes. Since the composites are constructed using eddies where the Sargassum presence is detected at least once during an eddy's lifetime, this could introduce a bias favoring instances of Sargassum being trapped inside eddies. To verify this, we performed a null hypothesis test by compositing the Sargassum distribution with the same criteria but using the eddy contours for the previous year. The results are shown in Fig. 4 and confirm that the eddy trapping observed in Fig. 3 and discussed below is not a bias of the methodology.
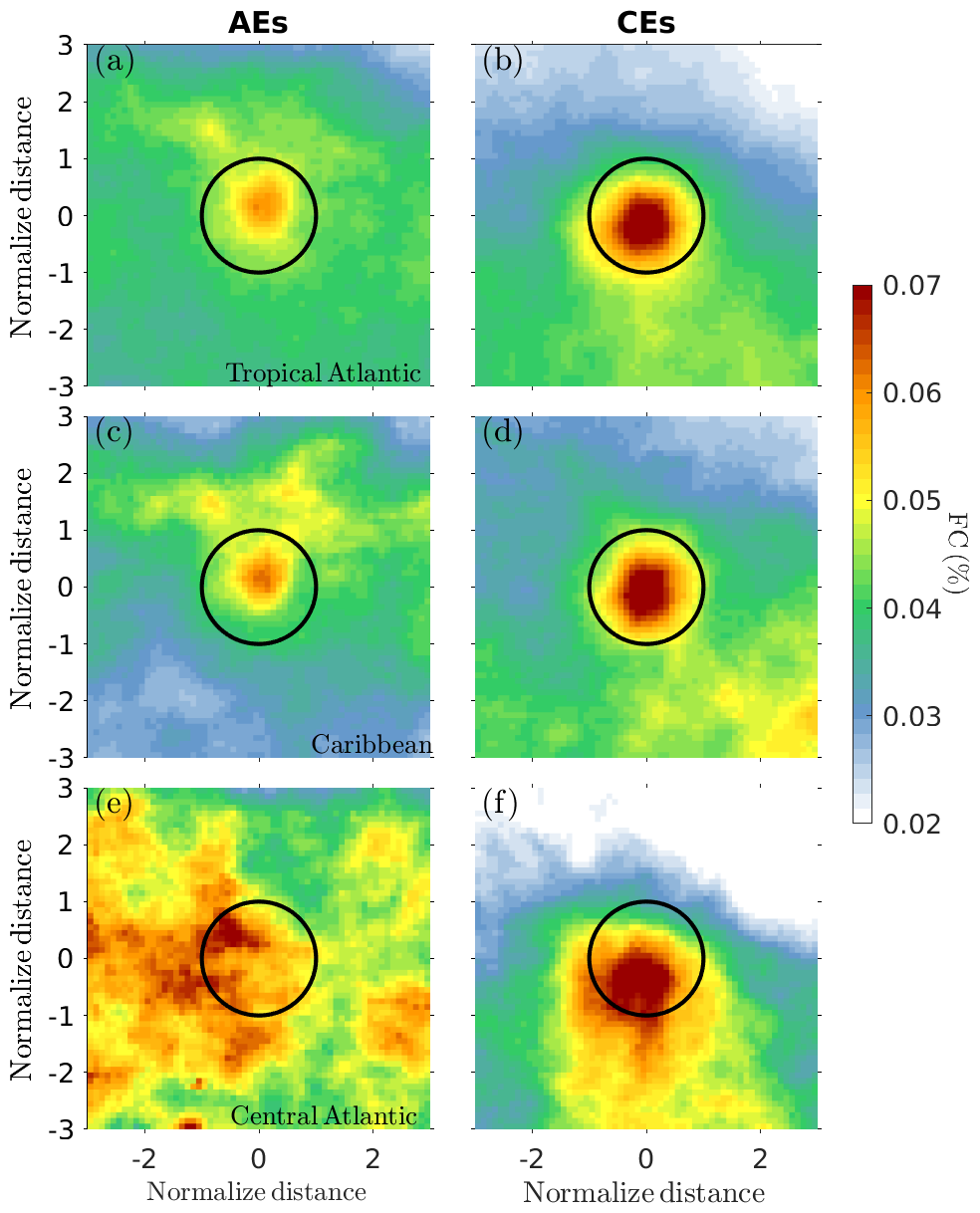
Figure 3Normalized composite averages of Sargassum cover (%; color) for AEs and CEs for the tropical Atlantic (a, b), Caribbean (c, d), and central Atlantic (e, f). The black circle represents an idealized contour of the eddy periphery.
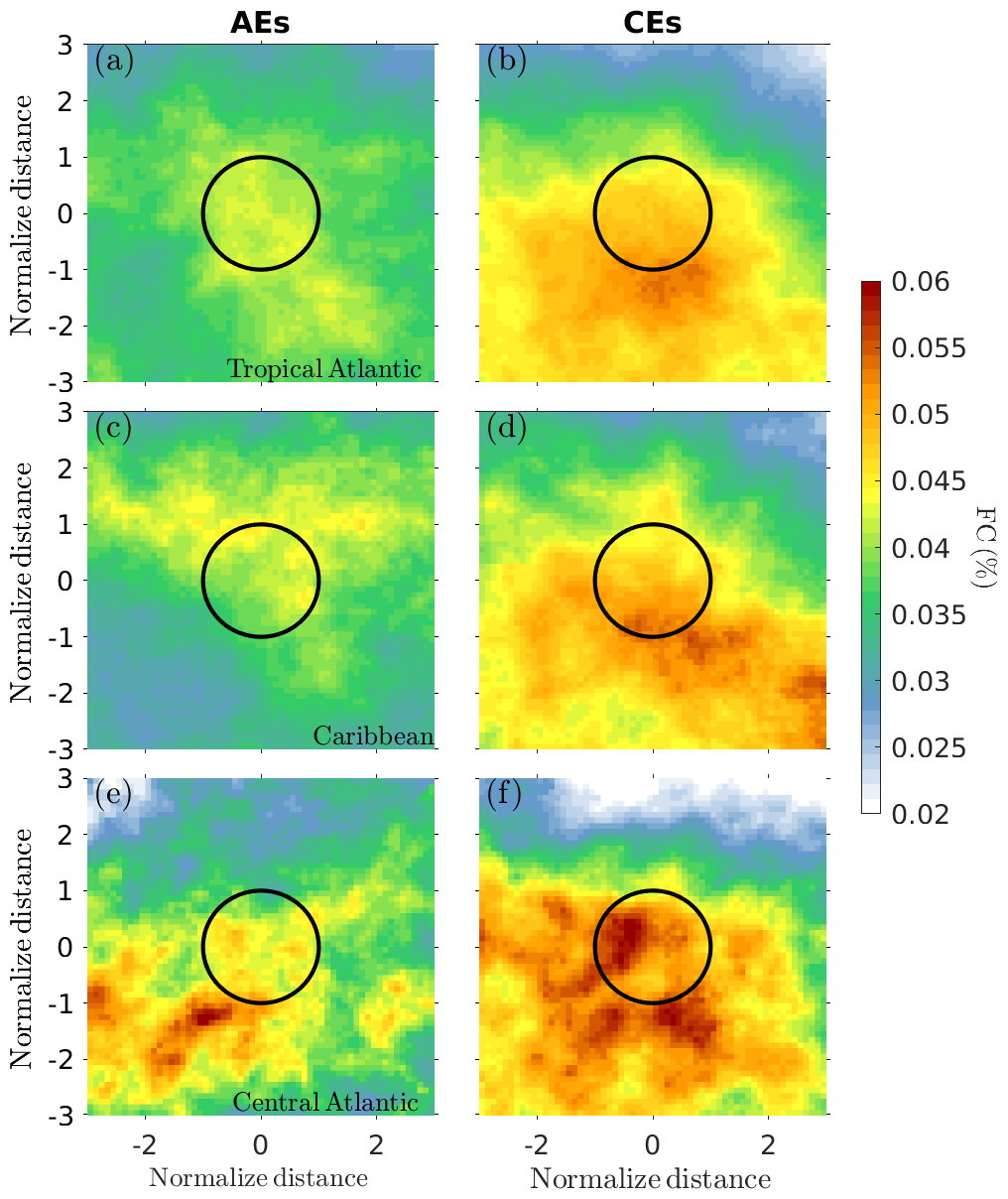
Figure 4Same as Fig. 3, but the Sargassum cover observed for a given day has been composited using eddy detections from the previous year. The same selection criteria were used as in Fig. 3. This is a null hypothesis that allows us to verify that the selection of the eddy cases for the compositing (e.g., that only eddies where Sargassum was observed are considered) does not induce a bias in favor of Sargassum accumulation inside the eddies.
The Sargassum cover composited for AEs and CEs in the tropical Atlantic is shown in Fig. 3a and b. It reveals that both anticyclones and cyclones accumulate Sargassum in their core but with much greater accumulation of Sargassum in cyclones (Fig. 3b) than in anticyclones (Fig. 3a). There are regional differences in this distribution. In the Caribbean, for example, the contrast between AEs and CEs is pronounced (Figs. 3c–d and 5d), whereas in the central Atlantic (Figs. 3e–f and 5g) it is much less pronounced, with a much more spread-out distribution for CEs and no evidence for accumulation in AEs. This could be due to the small number of AE samples in this region (Table 1). Also, note in Fig. 3b a meridional offset between the location of the maximum FC peak and the center of the eddies, unlike the Caribbean composites (Fig. 3d), where the Sargassum distribution appears centered within the eddies. The pattern observed in Fig. 3b may result from the influence of the central Atlantic cases included in the composite, which show a southward displacement of Sargassum relative to the eddy centers (see Fig. 3f). The cause of this southward shift is not fully understood at this stage, but it may be linked to windage effects from the trade winds or the background distribution of Sargassum, which generally tends to accumulate in the Intertropical Convergence Zone (ITCZ). Figure 4 also shows this latitudinal gradient of FC (see Fig. 4b and d), which means that this gradient is a result of sampling this peculiar region with this eddy scale.
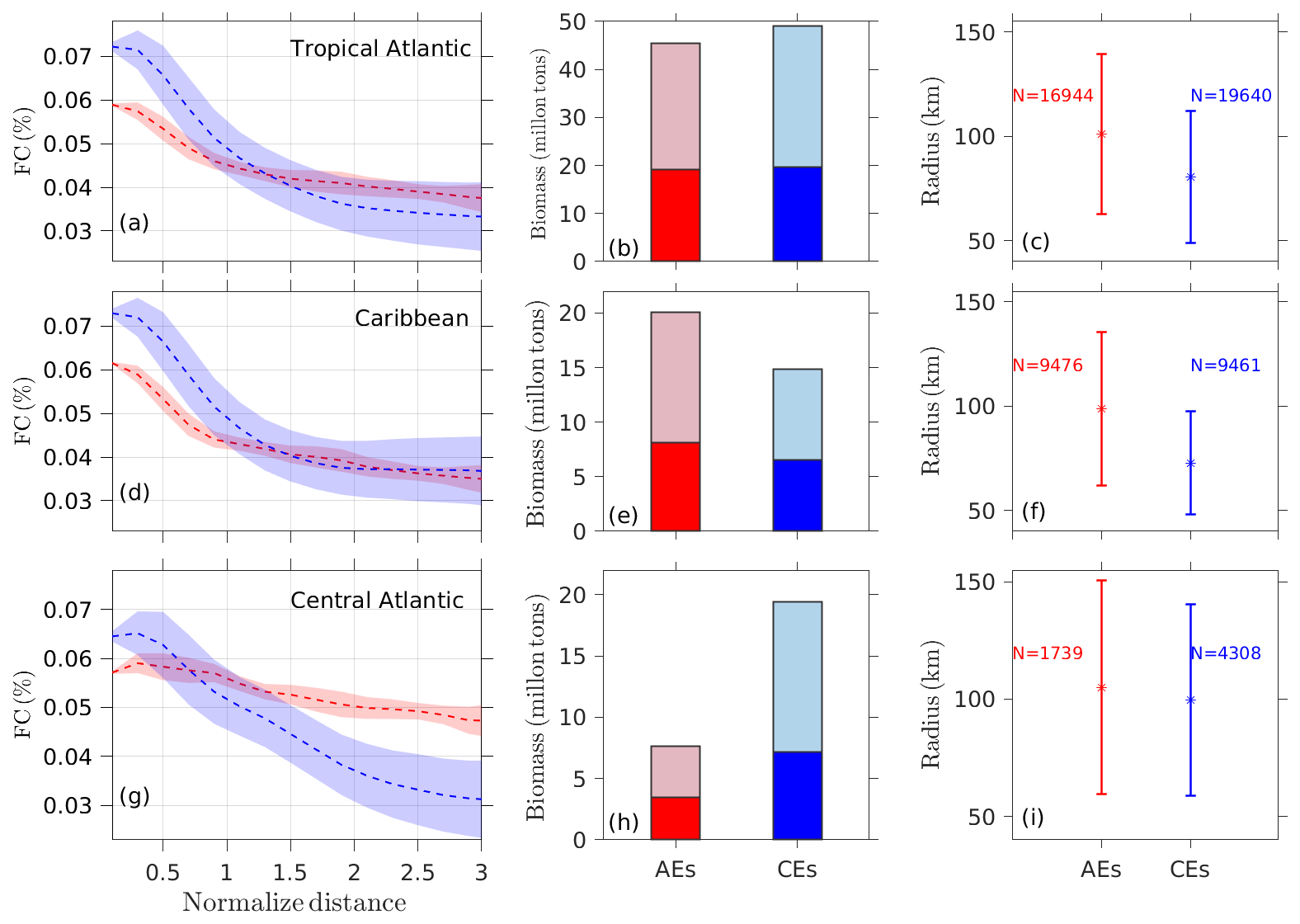
Figure 5(a, d, g) Sargassum cover averaged within concentric radii (represented as the normalized distance): the innermost radius was R=0.1 with an interval of 0.2 before reaching radius = 3 (the red and blue lines correspond to AE and CE, respectively), and the red and blue shadings represent 1 standard deviation of AE and CE. (b, e, h) Sargassum biomass within the AEs and CEs (light-red and blue bars) compared with the Sargassum biomass in eddies propagating for at least 30 d (dark-red and blue bars). (c, f, i) Radius distribution (±1 SD – standard deviation) of all eddies that contributed to building the composite, where N represents the number of eddies used for the calculation of the composites.
In terms of biomass, the AEs and CEs transport similar amounts of Sargassum at the basin scale (Fig. 5b), but again with significant regional differences (Fig. 5e–h). In the Caribbean, despite greater accumulation of Sargassum by cyclonic eddies, the anticyclones contain more Sargassum on average. This is explained by the difference in size between the two types of eddies in this region, with anticyclones being much larger than cyclones (Fig. 5f). In the central Atlantic, there are many more cyclonic eddies (Fig. 5i), which favors a higher biomass in cyclonic eddies. To quantify these differences in Table 1, we show the ratio of the total area and the biomass ratio of CEs vs. AEs. In the Caribbean, while the area ratio CEs AEs is 0.54, the accumulation of Sargassum in CEs leads to a biomass ratio CEs AEs of 0.80. These regional differences are illustrated well in Fig. 6a and b, which shows a large Sargassum content in the AEs of the North Brazil Current, the NBC ring pathways, and the northern Caribbean. The high Sargassum content transported by cyclonic eddies around 10° N corresponds to the shear zone between the North Equatorial Current and the North Equatorial Countercurrent (Fig. 6a and b).
Table 1Statistics of the Sargassum distribution within eddies in the Caribbean Sea and the central Atlantic.
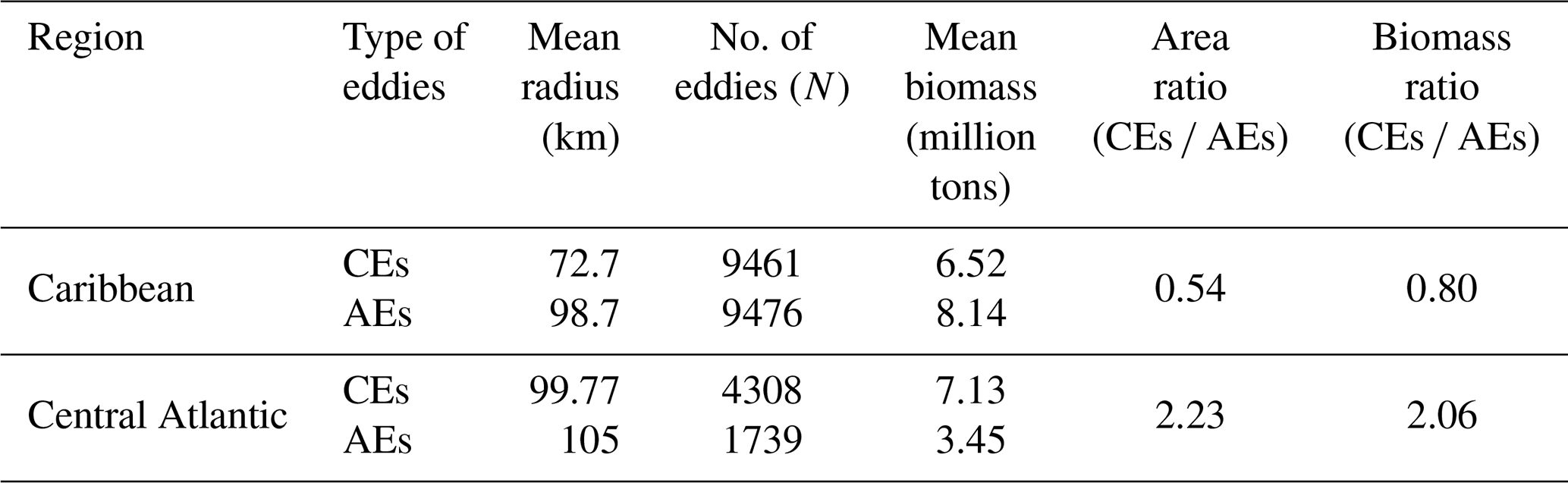
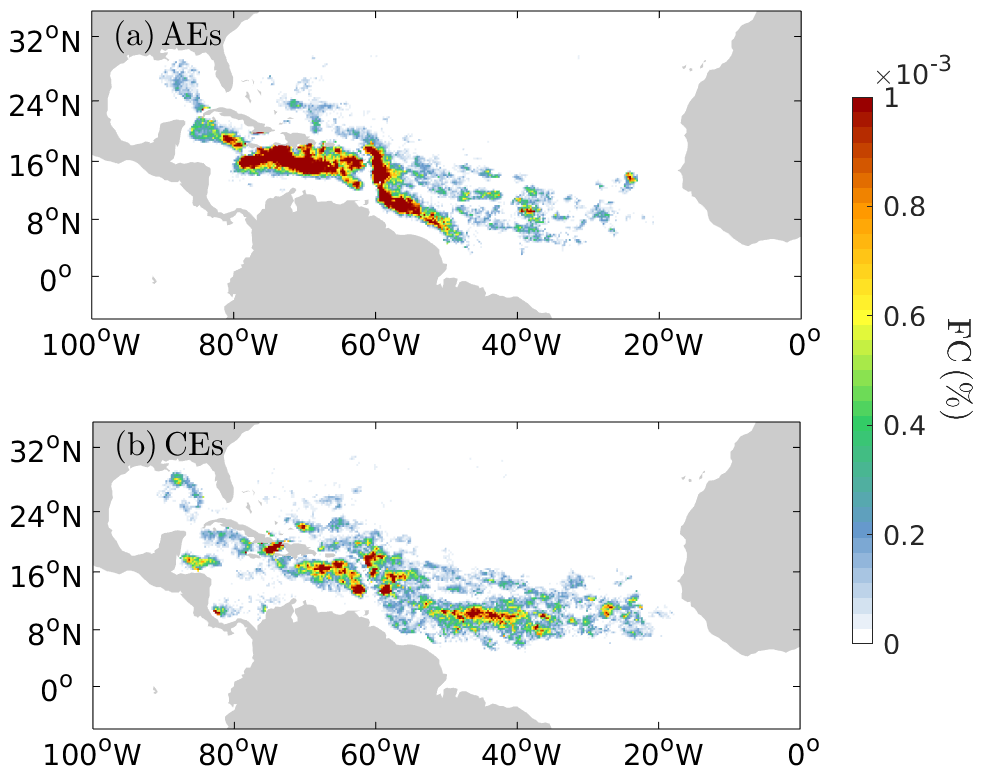
Figure 6The Sargassum cover (%) within AEs and CEs during the period from 2011 to 2023.
The distribution of the Sargassum varies in the eddy life cycle, as revealed by the evolution of the Sargassum cover with respect to the normalized lifetime of the eddies (Fig. 7). It shows that CEs increase their Sargassum cover along time for all of the regions considered, while the Sargassum cover within AEs shows a tendency to decrease along time, especially for the Caribbean Sea. The increase in the Sargassum cover at the end of the lifetime of the CEs in the central Atlantic could be explained by Ekman pumping. This mechanism depends on wind stress (trade winds across this region) and eddy velocity (Gaube et al., 2014), allowing convergence (divergence) of Ekman transport in CEs (AEs). This mechanism could favor the concentration of Sargassum in CEs. It is worth noting that the occurrence of AEs in the central Atlantic is low (Fig. 2) and may therefore contribute to a much noisier signal than in the Caribbean (Fig. 7b).
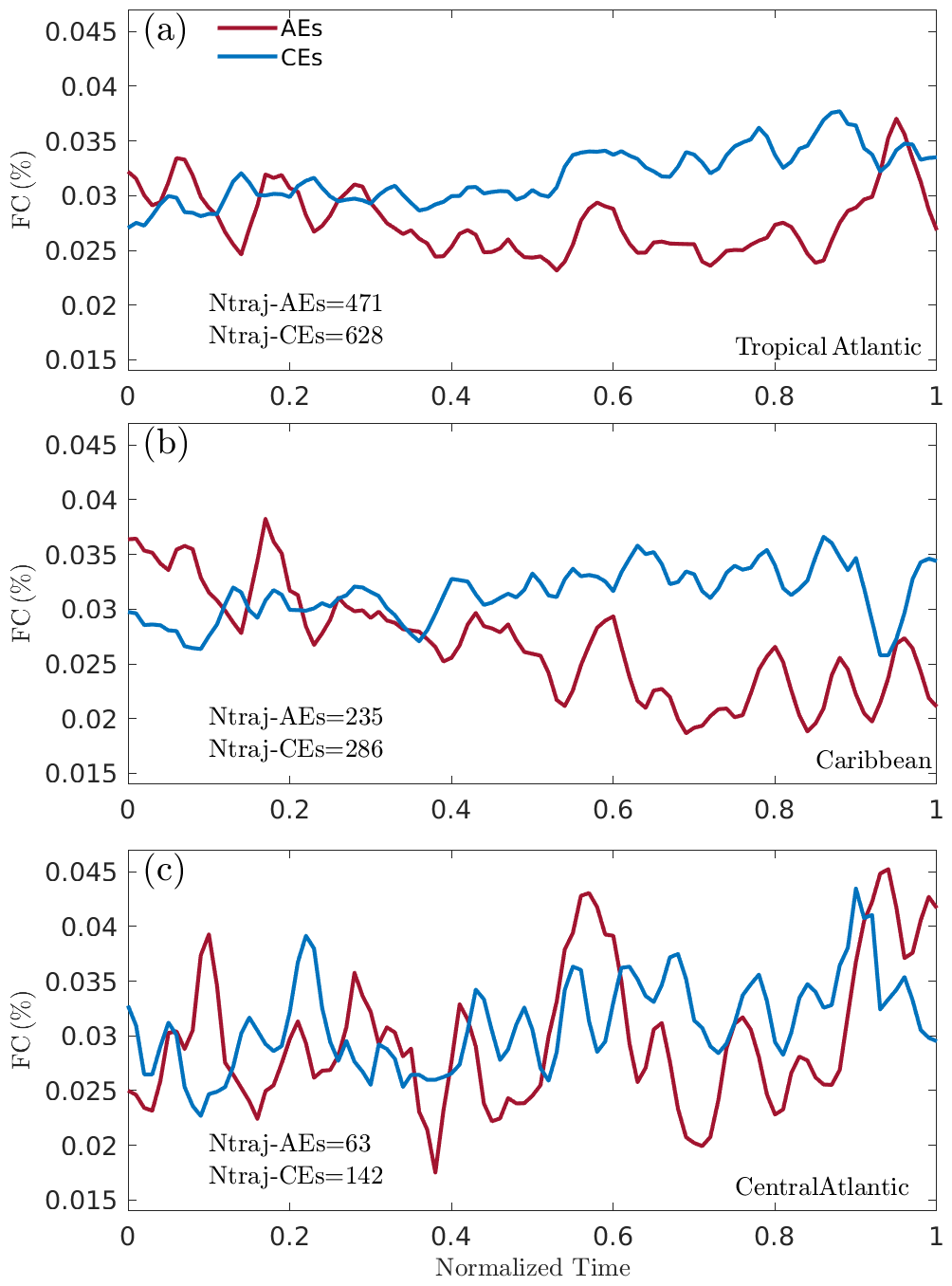
Figure 7(a) Average temporal evolution of Sargassum cover as a function of the normalized eddy lifetime (AEs and CEs, indicated by the red and blue lines, respectively). Ntraj-AEs and Ntraj-CEs are the number of trajectories considered for the different regions. (b) Caribbean and (c) central Atlantic.
Figure 8a shows the mean monthly biomass of Sargassum aggregated across the tropical Atlantic (5–30° N, 0–100° W), showing a clear seasonal cycle characterized by a growth phase from January to July, followed by a declining phase from September to December. In addition, estimates of Sargassum biomass within the AEs and CEs are shown (Fig. 8a). This provides insight into the periods when the retention of Sargassum biomass within the eddies was significant. Figure 8b illustrates the monthly percentage of Sargassum biomass contained within AEs and CEs relative to the total biomass. The accumulation of Sargassum biomass by these eddies can be significant, reaching up to 20 % in certain months. For example, in June 2018, this amount was equivalent to an estimated biomass of approximately 3 Mt (million tons) of Sargassum.
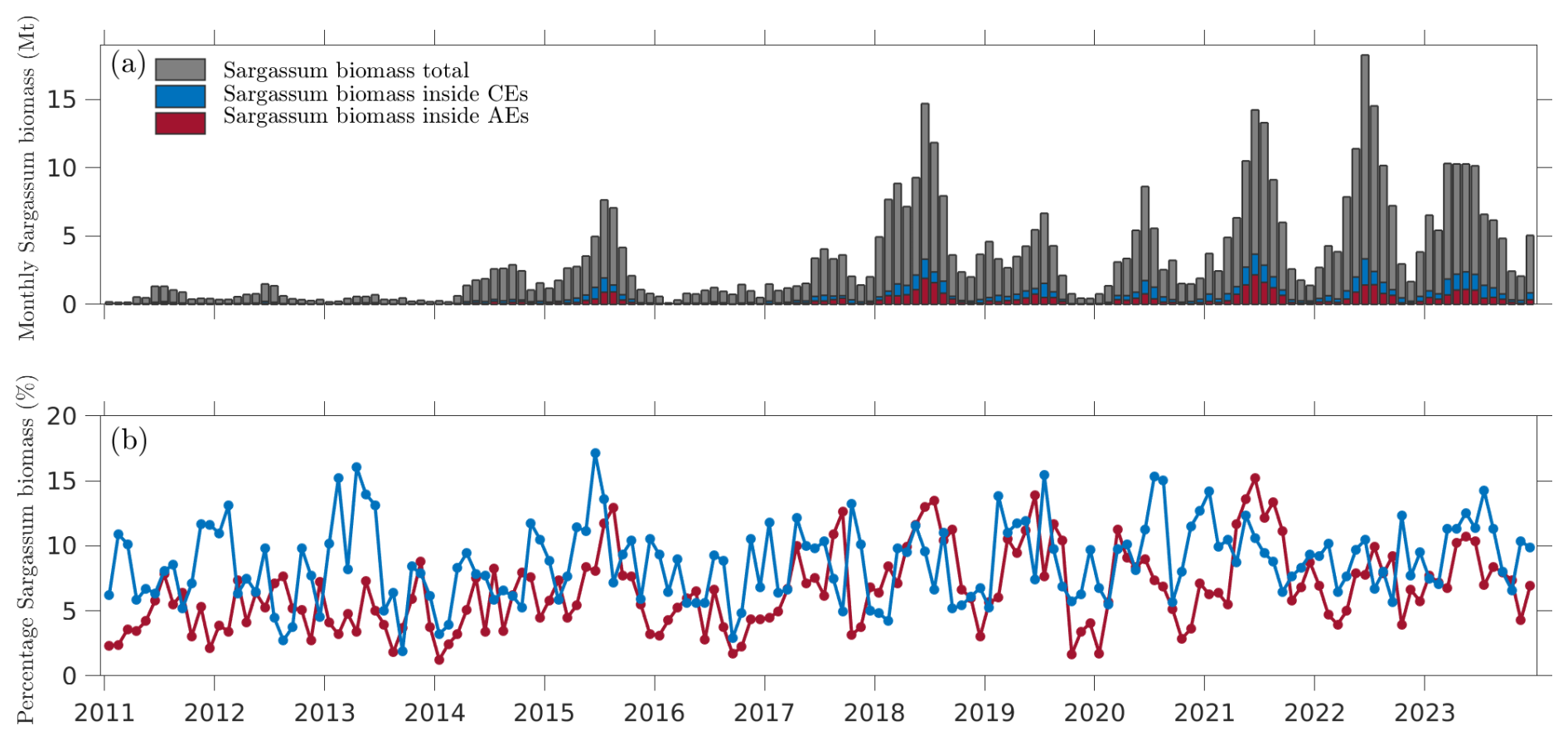
Figure 8(a) The total monthly biomass of Sargassum in the tropical Atlantic (gray bars, 5–30° N, 0–100° W) and within the AEs and CEs (red and blue bars in stacked form). (b) The monthly percentage of Sargassum biomass retained within the AEs and CEs relative to the total biomass (indicated by the red and blue lines, respectively).
This research provides new insights into the important role played by oceanic mesoscale eddies in structuring Sargassum biomass in the tropical Atlantic and fills a gap in our understanding of Sargassum organization by the ocean dynamical continuum, between the fine-scale aggregation processes (Langmuir, 1938; Zhong et al., 2012) and the large-scale distribution (Wang et al., 2019; Jouanno et al., 2023).
Using ADT satellite imagery, the distribution of Sargassum detections from 2011 to 2023 was analyzed by applying an eddy detection and tracking methodology (Chaigneau et al., 2009; Pegliasco et al., 2015). By combining these observational datasets, we showed that both cyclonic mesoscale and anticyclonic mesoscale eddies can trap and transport Sargassum. This suggests that these eddies serve as effective Sargassum transporters. The eddy detection method used in this study (Chaigneau et al., 2009) has been applied widely in the literature (e.g., Pegliasco et al., 2021; Ernst et al., 2023). Like any method, however, it has certain limitations. Its performance can be influenced by the resolution of the satellite data and the specific detection criteria employed. It is also important to exercise caution when interpreting Eulerian mesoscale eddy detections, as these rely on streamline-based approaches that are inherently observer-dependent (Andrade-Canto and Beron-Vera, 2022). In contrast, Lagrangian methods, such as geodesic eddy detection, identify eddies as coherent material structures that resist stretching and deformation over time and are therefore better suited to capturing flow-invariant transport pathways. In this work, we opted for an Eulerian detection method due to its relative ease of implementation. While it would be possible to reproduce this analysis using the methodology of Andrade-Canto and Beron-Vera (2022), we do not expect fundamentally different results, particularly because our detection method does not identify eddies that cross the Antilles Arc or the Yucatan Channel (see Fig. 2), as was the case in Huang et al. (2021). That said, Lagrangian approaches would be particularly valuable for investigating transport and accumulation mechanisms, especially in scenarios where eddies reform in the wake of islands.
Our composite analysis of Sargassum cover within mesoscale eddies consistently shows a preference for Sargassum accumulation within CEs over AEs in the tropical Atlantic, with an average of 15 % higher Sargassum cover in CEs. Although a drogued drifter dataset is used, this is consistent with recent findings by Vic et al. (2022), who showed a 24 % higher accumulation of drifters in CEs compared to AEs in the North Atlantic using observed and simulated trajectories. The tendency to accumulate or not in cyclonic and anticyclonic eddies has been shown to depend on theoretical choices and the complexity of drift models (Provenzale, 1999; Beron-Vera, 2021), so our findings should have important implications for motivating future drift model developments.
The results also suggest that the trapping is much more effective in the Caribbean than in the central Atlantic, as revealed by the sharpest contrast of Sargassum cover between the interior and exterior in the Caribbean. One hypothesis is that there is much less energetic mesoscale activity in the central Atlantic compared to the Caribbean: weak eddy circulation and the drift associated with the trade winds may reduce the capacity of the eddies to accumulate Sargassum.
These results raise the question of how mesoscale activity could influence the growth and decay of Sargassum by modulating nutrient availability. It has been documented that processes such as Ekman pumping and eddy pumping act by increasing nutrients in the euphotic layer in cyclones and decreasing nutrients in the euphotic layer in anticyclones (Gaube et al., 2014; McGillicuddy, 2016). In addition to contrasted surface radial velocity properties between AEs and CEs, this could contribute to the contrasted time evolution of the distribution highlighted in Fig. 7 and could modulate the overall growth of the total Sargassum biomass at the basin scale. This should receive further attention in the future.
All of the code and processed data needed to reproduce the main results and figures in this paper have been made available via Zenodo (https://doi.org/10.5281/zenodo.14816717, Sosa-Gutierrez et al., 2025).
RSG, JJ, and LB conceived the study. RSG and JJ wrote the manuscript. RSG analyzed the data. JJ acquired the funding and managed the project. All of the authors reviewed and edited the draft version.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
The Sargassum cover database was processed by the AERIS/ICARE Data and Services Center at the University of Lille in collaboration with MIO (https://doi.org/10.12770/8fe1cdcb-f4ea-4c81-8543-50f0b39b4eca). Finally, we would like to thank the reviewers Maria Josefina Olascoaga and Clément Vic for their valuable comments that helped improve this article. Open research: the remote sensing ADT observations are available from the Global Ocean Gridded L4 Sea Surface Heights And Derived Variables Reprocessed 1993 Ongoing Copernicus Marine Service, and theSargassum fractional cover is available at https://www.odatis-ocean.fr (last access: June 2024).
This research has been supported by the Agence Nationale de la Recherche (grant no. ANR-19-SARG-0007) and CNES under projects TOSCA-SAREDA-HR and TOSCA-SARGAT.
This paper was edited by Erik van Sebille and reviewed by M. Josefina Olascoaga and Clément Vic.
Andrade-Canto, F. and Beron-Vera, F. J.: Do eddies connect the tropical Atlantic Ocean and the Gulf of Mexico?, Geophys. Res. Lett., 49, e2022GL099637, https://doi.org/10.1029/2022GL099637, 2022.
Andrade-Canto, F., Beron-Vera, F. J., Goni, G. J., Karrasch, D., Olascoaga, M. J., and Triñanes, J.: Carriers of Sargassum and mechanism for coastal inundation in the Caribbean Sea, Phys. Fluids, 34, 016602, https://doi.org/10.1063/5.0079055, 2022.
Antonio-Martínez, F., Henaut, Y., Vega-Zepeda, A., Cerón-Flores, A. I., Raigoza-Figueras, R., Cetz-Navarro, N. P., and Espinoza-Avalos, J.: Leachate effects of pelagic Sargassum spp. on larval swimming behavior of the coral Acropora palmata, Sci. Rep., 10, 3910, https://doi.org/10.1038/s41598-020-60864-z, 2020.
Beron-Vera, F. J.: Nonlinear dynamics of inertial particles in the ocean: From drifters and floats to marine debris and Sargassum, Nonlin. Dynam., 103, 1–26, 2021.
Beron-Vera, F. J., Olascoaga, M. J., Haller, G., Farazmand, M., Triñanes, J., and Wang, Y.: Dissipative inertial transport patterns near coherent Lagrangian eddies in the ocean, Chaos, 25, 087412, https://doi.org/10.1063/1.4928693, 2015.
Brach, L., Deixonne, P., Bernard, M.-F., Durand, E., Desjean, M.-C., Perez, E., van Sebille, E., and ter Halle, A.: Anticyclonic eddies increase accumulation of microplastic in the North Atlantic subtropical gyre, Mar. Pollut. Bull., 126, 191–196, https://doi.org/10.1016/j.marpolbul.2017.10.077, 2018.
Chaigneau, A., Eldin, G., and Dewitte, B.: Eddy activity in the four major upwelling systems from satellite altimetry, (1992–2007), Prog. Oceanogr., 83, 117–123, https://doi.org/10.1016/j.pocean.2009.07.012, 2009.
Chen, G., Hou, Y., and Chu, X.: Mesoscale eddies in the South China Sea: Mean properties, spatiotemporal variability, and impact on thermohaline structure, J. Geophys. Res., 116, C06018, https://doi.org/10.1029/2010JC006716, 2011.
D'Asaro, E. A., Shcherbina, A. Y., Klymak, J. M., Molemaker, J., Novelli, G., Guigand, C. M., Haza, A. C., Haus, B. K., Ryan, E. H., Jacobs, G. A., Huntley, H. S., Laxague, N. J. M., Chen, S., Judt, F., McWilliams, J. C., Barkan, R., Kirwan, A. D., Poje, A. C., and Özgökmen, T. M.: Ocean convergence and the dispersion of flotsam, P. Natl. Acad. Sci. USA, 115, 1162–1167, https://doi.org/10.1073/pnas.1718453115, 2018.
Descloitres, J., Minghelli, A., Steinmetz, F., Chevalier, C., Chami, M., and Berline, L.: Revisited Estimation of Moderate Resolution Sargassum Fractional Coverage Using Decametric Satellite Data (S2/MSI), Remote Sens., 13, 5106, https://doi.org/10.3390/rs13245106, 2021.
Ernst, P. A., Subrahmanyam, B., Trott, C. B., and Chaigneau, A.: Characteristics of submesoscale eddy structures within mesoscale eddies in the Gulf of Mexico from 1/48° ECCO estimates, Front. Mar. Sci., 10, 1181676, https://doi.org/10.3389/fmars.2023.1181676, 2023.
Esposito, G., Berta, M., Centurioni, L., Lodise, J., Ozgokmen, T., Poulain, P.-M., and Griffa, A.: Submesoscale vorticity and divergence in the Alboran Sea: Scale and depth dependence, Front. Mar. Sci., 8, 843, https://doi.org/10.3389/fmars.2021.678304, 2021.
EU Copernicus Marine Service Product: Global Ocean Gridded L 4 Sea Surface Heights And Derived Variables Reprocessed Copernicus Climate Service, Mercator Ocean International [data set], https://doi.org/10.48670/moi-00145, 2024.
Gaube, P., McGillicuddy, D. J., Chelton, D. B., Behrenfeld, M. J., and Strutton, P. G.: Regional variations in the influence of mesoscale eddies on near-surface chlorophyll, J. Geophys. Res.-Oceans, 119, 8195–8220, https://doi.org/10.1002/2014JC010111, 2014.
Gower, J. F. R. and King, S. A.: Distribution of floating Sargassum in the Gulf of Mexico and the Atlantic Ocean mapped using MERIS, Int. J. Remote Sens., 32, 1917–1929, https://doi.org/10.1080/01431161003639660, 2011.
Gower, J., Young, E., and King, S.: Satellite images suggest a new Sargassum source region in 2011, Remote Sens. Lett., 4, 764–773, 2013.
Hendy, I. W., Woolford, K., Vincent-Piper, A., Burt, O., Schaefer, M., Cragg, S. M., Sanchez-Navarro, P., and Ragazzola, F.: Climate-driven golden tides are reshaping coastal communities in Quintana Roo, Mexico, Clim. Change Ecol., 2, 100033, https://doi.org/10.1016/j.ecochg.2021.100033, 2021.
Huang, M., Liang, X., Zhu, Y., Liu, Y., and Weisberg, R. H.: Eddies connect the tropical Atlantic Ocean and the Gulf of Mexico, Geophys. Res. Lett., 48, e2020GL091277, https://doi.org/10.1029/2020GL091277, 2021.
Jouanno, J., Sheinbaum, J., Barnier, B., Molines, J.-M., Debreu, L., and Lemarié, F.: The mesoscale variability in the caribbean sea. part i: Simulations and characteristics with an embedded model, Ocean Model., 23, 82–101, https://doi.org/10.1016/j.ocemod.2008.04.002, 2008.
Jouanno, J., Morvan, G., Berline, L., Benshila, R., Aumont, O., Sheinbaum, J., and Ménard, F.: Skillful seasonal forecast of Sargassum proliferation in the Tropical Atlantic, Geophys. Res. Lett., 50, e2023GL105545, https://doi.org/10.1029/2023GL105545, 2023.
Langmuir, I.: Surface motion of water induced by wind, Science, 87, 119–123, 1938.
Lévy, M., Franks, P. J., and Smith, K. S.: The role of submesoscale currents in structuring marine ecosystems, Nat. Commun., 9, 4758, https://doi.org/10.1038/s41467-018-07059-3, 2018.
McGillicuddy Jr., D. J.: Mechanisms of physical-biological-biogeochemical interaction at the oceanic mesoscale, Annu. Rev. Mar. Sci., 8, 125–159, https://doi.org/10.1146/annurev-marine-010814-015606, 2016.
Nencioli, F., Dong, C., Dickey, T., Washburn, L., and McWilliams, J. C.: A vector geometry-based eddy detection algorithm and its application to a high-resolution numerical model product and high-frequency radar surface velocities in the Southern California Bight, J. Atmos. Ocean. Tech., 27, 564–579, https://doi.org/10.1175/2009JTECHO725.1, 2010.
Ody, A., Thibaut, T., Berline, L., Changeux, T., André, J.-M., Chevalier, C., Blanfuné, A., Blanchot, J., Ruitton, S., Stiger-Pouvreau, V., Connan, S., Grelet, J., Aurelle, D., Guéné, M., Bataille, H., Bachelier, C., Guillemain, D., Schmidt, N., Fauvelle, V., Guasco, S., and Ménard, F.: From in situ to satellite observations of pelagic Sargassum distribution and aggregation in the Tropical North Atlantic Ocean, Plos ONE, 14, e0222584, https://doi.org/10.1371/journal.pone.0222584, 2019.
Okubo, A.: Horizontal dispersion of floatable particles in vicinity of velocity singularities such as convergences, Deep-Sea Res., 17, 445–454, https://doi.org/10.1016/0011-7471(70)90059-8, 1970.
Pegliasco, C., Chaigneau, A., and Morrow, R.: Main eddy vertical structures observed in the four major Eastern Boundary Upwelling Systems, J. Geophys. Res.-Oceans, 120, 6008–6033, 2015.
Pegliasco, C., Chaigneau, A., Morrow, R., and Dumas, F.: Detection and tracking of mesoscale eddies in the Mediterranean Sea: A comparison between the Sea Level Anomaly and the Absolute Dynamic Topography fields, Adv. Space Res., 68, 401–419, https://doi.org/10.1016/j.asr.2020.03.039, 2021.
Provenzale, A.: Transport by coherent barotropic vortices, Annu. Rev. Fluid Mech., 31, 55–93, https://doi.org/10.1146/annurev.fluid.31.1.55, 1999.
Richardson, P. L.: Caribbean Current and eddies as observed by surface drifters, Deep-Sea Res. Pt. II, 52, 429–463, 2005.
Rodríguez-Martínez, R. E., Reali, M. Á. G., Torres-Conde, E. G., and Bates, M. N.: Temporal and spatial variation in hydrogen sulfide (H2S) emissions during holopelagic Sargassum spp. decomposition on beaches, Environ. Res., 247, 118235, https://doi.org/10.1016/j.envres.2024.118235, 2024.
Rosellón-Druker, J., McAdam-Otto, L., Suca, J. J., Seary, R., Gaytán-Caballero, A., Escobar-Briones, E., Hazen, E., and Muller-Karger, F.: Local ecological knowledge and perception of the causes, impacts and effects of Sargassum massive influxes: a binational approach, Ecosyst. People, 19, 2253317, https://doi.org/10.1080/26395916.2023.2253317, 2023.
Sosa-Gutiérrez, R., Pallàs-Sanz, E., Jouanno, J., Chaigneau, A., Candela, J., and Tenreiro, M.: Erosion of the subsurface salinity maximum of the loop current eddies from glider observations and a numerical model, J. Geophys. Res.-Oceans, 125, e2019JC015397, https://doi.org/10.1029/2019JC015397, 2020.
Sosa-Gutierrez, R., Jouanno, J., Berline, L., Descloitres, J., and Chevalier, C.: Impact of tropical cyclones on pelagic Sargassum, Geophys. Res. Lett., 49, e2021GL097484, https://doi.org/10.1029/2021GL097484, 2022.
Sosa-Gutierrez, R., Jouanno, J., and Berline, L.: Sargassum accumulation and transport by mesoscale eddies [Data set], Zenodo [data set], https://doi.org/10.5281/zenodo.14816717, 2025.
Sun, Y., Wang, M., Liu, M., Li, Z. B., Chen, Z., and Huang, B.: Continuous Sargassum monitoring across the Caribbean Sea and Central Atlantic using multi-sensor satellite observations, Remote Sens. Environ., 309, 114223, https://doi.org/10.1016/j.rse.2024.114223, 2024.
Van Tussenbroek, B. I., Arana, H. A. H., Rodríguez-Martínez, R. E., Espinoza-Avalos, J., Canizales-Flores, H. M., González-Godoy, C. E., Barba-Santos, M. G., Vega-Zepeda, A., and Collado-Vides, L.: Severe impacts of brown tides caused by Sargassum spp. on near-shore Caribbean seagrass communities, Mar. Pollut. Bull., 122, 272–281, https://doi.org/10.1016/j.marpolbul.2017.06.057, 2017.
Vic, C., Hascoët, S., Gula, J., Huck, T., and Maes, C.: Oceanic mesoscale cyclones cluster surface Lagrangian material, Geophys. Res. Lett., 49, e2021GL097488, https://doi.org/10.1029/2021GL097488, 2022.
Wang, M. and Hu, C.: Mapping and quantifying Sargassum distribution and coverage in the Central Western Atlantic using MODIS observations, Remote Sens. Environ., 183, 350–367, https://doi.org/10.1016/j.rse.2016.04.019, 2016.
Wang, M., Hu, C., Cannizzaro, J., English, D., Han, X., Naar, D., Lapointe, B., Brewton, R., and Hernandez, F.: Remote sensing of Sargassum biomass, nutrients, and pigments, Geophys. Res. Lett., 45, 12359–12367, 2018.
Wang, M., Hu, C., Barnes, B. B., Mitchum, G., Lapointe, B., and Montoya, J. P.: The great Atlantic Sargassum belt, Science, 365, 83–87, https://doi.org/10.1126/science.aaw7912, 2019.
Weiss, J.: The dynamics of enstrophy transfer in 2-dimensional hydrodynamics, Physica D, 48, 273–294, https://doi.org/10.1016/0167-2789(91)90088-Q, 1991.
Zhong, Y., Bracco, A., and Villareal, T. A.: Pattern formation at the ocean surface: Sargassum distribution and the role of the eddy field, Limnol. Oceanogr.: Fluids Environ., 2, 12–27, https://doi.org/10.1215/21573689-1573372, 2012.